|
Читайте также: |
|
|
 Р* Р
Р* Р
В этом уравнении k – суммарная константа скорости всех процессов, в которых расходуются (инактивируются) возбужденные молекулы.
Согласно правилам химической кинетики, изменение концентрации возбужденных молекул в образце (d[P*]) за время dt будет равно:
|
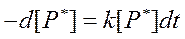
Решим полученное дифференциальное уравнение (67):
|
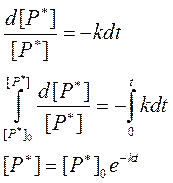
В представленных выражениях [P*]0 – концентрация возбужденных молекул в момент прекращения их генерации (момент времени t =0). Из уравнения (68) следует, что после прекращения генерации новых возбужденных молекул их концентрация в объекте будет экспоненциально снижаться со временем.
Константа скорости инактивации возбужденных молекул k имеет размерность 1/с. Введем величину t = 1/k. Обозначим ее как реальное время жизни возбужденных состояний в образце и выразим с ее помощью зависимость [P*] от времени, преобразовав выражение (68):
|
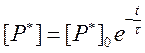
Определим, насколько измениться [P*] к моменту времени t=t:
|
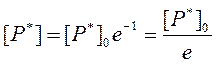
Из уравнения (70) следует, таким образом, что к моменту времени t=t концентрация возбужденных молекул в образце после прекращения их генерации de novo снизится относительно исходной концентрации в е раз (е – основание натуральных логарифмов, равное 2,7182818284 5904523536…).
Константа скорости инактивации возбужденных молекул k вводилась нами, как суммарная константа скоростей всех процессов, в ходе которых эти молекулы расходуются. Иными словами, k=k1+k2+k3+…+kn, где k1, k2, k3 и т.д. - константы скорости каждого из процессов, сопровождающихся исчезновением молекул в состоянии возбуждения в объекте. Пусть при этом k1 – константа скорости излучательного тушения таких молекул:
|
|
Тогда можно выразить интенсивности фотолюминесценции образца через концентрации возбужденных молекул в нем следующим образом:
|

|
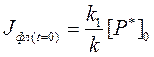
Подставив выражение [P*] и [P*]0 через соответствующие значения Jфл из уравнений (72) и (73), в уравнение (69), получим
|

Иными словами, интенсивность фотолюминесценции объекта после прекращения его возбуждения также, как и концентрация возбужденных молекул в образце, будет снижаться со временем экспоненциально. Это дает возможность определения величины t (реального времени жизни возбужденных состояний в образце) экспериментально. Для определения t нужно зарегистрировать кинетику затухания фотолюминесценции после выключения источника возбуждающего света, и определить время, за которое Jфл снижается в е раз. Следует заметить, что это сделать бывает непросто – напомним, что синглетно-возбужденные молекулы при обычных условиях существуют не более 10-12-10-8 с. Поэтому для регистрации кинетик затухания фотолюминесценции требуются чрезвычайно быстродействующие приборы, с константой времени реагирования порядка 10-13 с. Впрочем, в последнее время для этого применяются специальные устройства – однофотонные t -метры (читается – тау-метры). В этих приборах регистрируется распределение по величине времени жизни отдельных молекул в объекте после возбуждения фотолюминесценции коротким (продолжительностью от 0,1 до 10 нс) импульсом излучения. Затем по этому распределению рассчитывается искомая величина t.
Выше уже неоднократно указывалось, что испускание кванта фотолюминесценции в обычных условиях – не единственный способ избавления от энергии электронного возбуждения для возбужденных молекул. Однако можно представить себе условия, в которых фотолюминесценция будет таковым. Действительно, если возбужденная молекула находится при температуре около абсолютного нуля и в вакууме, т.е. всякие ее взаимодействия с окружением исключены, то величина k1 для нее будет стремиться к k. Квантовый же выход фотолюминесценции в таких условиях станет равен 1. Если рассматривать время жизни возбужденных молекул в таких, модельных, условиях, когда квантовый выход фотолюминесценции равен 1 (т.е. все возбужденные молекулы инактивируются исключительно излучательно), то этот показатель принято называть истинным временем жизни возбужденного состояния. Обозначим его tи. Введя понятие tи, легко составить пропорцию, показывающую взаимосвязь между реальным и истинным временами жизни возбужденных состояний, с одной стороны, и квантовым выходом фотолюминесценции – с другой.
Действительно, если величине tи соответствует квантовый выход фотолюминесценции 1, а величине t - квантовый выход фотолюминесценции q, то:
|
t=tи q
а
|
Выражения (75) и (76) определяют, таким образом, как связаны квантовый выход фотолюминесценции и времена жизни возбужденных состояний молекул в объекте друг с другом.
Если мы вспомним, что понятие времени жизни возбужденного состояния вводилось нами, как величина, обратная константе скорости инактивации электронных возбуждений (т.е. t=1/k, а tи=1/k1, см. выше), можно также написать, что:
|
Следует заметить, что измерение величин t обычно осуществляется для определения типа электронного возбуждения исследуемых молекул в объекте. Это бывает необходимо, например, при выяснении того, какая именно молекула в объекте может опосредовать фотохимические реакции, определяющие тот или иной фотобиологический эффект.
Как уже указывалось выше, при триплетном возбуждении обратный переход Т ® S0 требует от электрона изменения спина, т.е. дополнительных затрат энергии. Поэтому триплетно-возбужденные молекулы живут в состоянии возбуждения намного дольше, чем синглетно-возбужденные (напомним, что для синглетных возбуждений время жизни составляет 10-8 ¸10-12 секунды, а для триплетных – от 10-7 секунды до целых секунд). Это высказывание относится, впрочем, только к величинам tи рассматриваемых типов возбужденных молекул. Дело здесь в том, что и у синглетных, и у триплетных возбужденных состояний время жизни существенно зависит от наличия возможности безизлучательного избавления от избытка свободной энергии. С ростом температуры такая возможность появляется (k1 перестает быть равным k, поскольку появляются процессы инактивации возбуждений с константами скоростей k2, k3 и т.д.). Поэтому t всегда оказывается меньше, чем tи. Однако, из-за того, что tи у триплетно-возбужденных молекул изначально больше, эти молекулы успевают за время жизни в возбужденном состоянии сместиться от места образования дальше (чем синглетно-возбужденные), и для них вероятность безизлучательного тушения (например, путем растраты энергии в ходе соударений с другими молекулами) с ростом температуры увеличивается значительно быстрее. Из-за этого, в частности, зарегистрировать фосфоресценцию большинства биологических объектов при комнатной температуре (порядка 300о К) из-за ее малой интенсивности очень сложно. Большая величина tи, кроме того, позволяет молекулам в состоянии триплетного возбуждения, в сравнении с синглетно-возбужденными, чаще вступать в фотохимические реакции. Поэтому многие фотобиологические процессы оказываются связаны с фотохимическим превращениями, опосредуемыми именно триплетно-возбужденными молекулами.
Важно заметить, что величина константы скорости излучательного перехода в возбужденных молекулах (k1) зависит от способности молекул данного типа поглощать излучение в основном состоянии:
|

В выражении (78) v – волновое число излучения (v=1/l), vм – величина волнового числа в максимуме поглощения, интеграл – площадь под кривой спектра поглощения (для одной полосы поглощения этот интеграл составляет» eмDv1/2, eм – величина молярного коэффициента поглощения в максимуме рассматриваемой полосы, Dv1/2 – ее полуширина). Из уравнения (78) вытекает, что величина tи падает, если возбуждение молекул осуществляется излучением в интенсивных максимумах их спектров поглощения.
Дата добавления: 2015-07-25; просмотров: 379 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Влияние светорассеивания на фотолюминесценцию. | | | Влияние окружения люминесцирующих молекул на параметры фотолюминесценции. Флуоресцентные зонды и метки. |