Читайте также:
|
Крайне сложно дифференцированная у человека топографическая карта разделения коры мозга на области и поля является результатом длительного прогрессивного развития первоначально слаборазвитой и в значительной мере еще слитно представленной нерасчлененной корковой организации. У низших млекопитающих могут быть выделены лишь немногие основные зоны анализаторов (зрительная, слуховая, кожно-кинестетическая), еще не дифференцированные внутри себя на тюля и очень неотчетливо разграниченные между собой (рис. 15). На более высоких ступенях эволюции млекопитающих в задней части полушария происходит все более отчетливое отграничение ядерных зон анализаторов с выделением в них центральных и периферических полей. Одновременно с этим процессом в связи с интенсивным ростом по поверхности полушария самих ядерных зон происходит все большее надвигание их одна на другую с образованием «зон перекрытия» (рис. 16). Последние еще слабо обособлены у хищных и копытных, отчетливо выделяясь, однако, у всех приматов. В пе редней части полушария прогрессивная дифференцировка корковой организации протекает в направлении выделения двигательной зоны коры, с последующим обособлением моторного и премоторного полей и интенсивным ростом территории, занимаемой полями собственно лобной области. Уже у человекообразных обезьян в последней выделяются почти все те поля, что и у человека; однако лишь у человека достигает своего завершения качественная дифференцировка структуры этих полей и устанавливаются их окончательные топографические соотношения.
У человека в огромной степени возрастает территория, занимаемая третичными полями, которые в совокупности составляют свыше половины всей протяженности коры. Одновременно с этим сильно возрастает также площадь периферических полей по сравнению с центральными полями, которые занимают у человека относительно небольшие участки (рис. 17), хотя по абсолютной своей величине и эти поля увеличиваются по протяжению по сравнению с такими же полями у обезьян.
Выделенные нами три группы полей формируются в определенной последовательности как на протяжении эволюции млекопитающих, так и в течение индивидуального развития организма. Центральные поля выделяются наиболее рано по сравнению со всеми остальными и отличаются особенностями своей цитоархитектоники и нейронного строения; эти отличия появляются как в филогенезе, так и в пренатальном онтогенезе. Уже фундаментальными исследованиями Флексига (1920, 1927), а затем Фогтов (1919—1920) и др. было показано, что и системы связей этих полей с подкорковыми образованиями мозга созревают и миэлинизируются первыми (рис. 18). Периферические поля ядерных зон заметно позднее, чем центральные поля, вступают в интенсивный период своего развития. В онтогенезе человека этот период падает на первые недели и месяцы жизни. Соответственно и процесс миэлинизации проводящих путей, связанных с этими полями, протекает в более поздние сроки. Наиболее поздно созревают зоны перекрытия анализаторов и формации лобной области; это наблюдается только на протяжении первых лет жизни ребенка. Связи этих корковых территорий также наиболее поздно завершают цикл своего развития.

Рис. 15. Прогрессивная дифференциация областей и полей коры головного мозга (по Г. И. Полякову): А — мозг ежа; Б — мозг крысы; В — мозг собаки; Г — мозг низшей обезьяны; Д — мозг высшей обезьяны; E — мозг человека. Крупными точками обозначены первичные (центральные) поля ядерных зон. Точками средней величины — периферические (вторичные) поля ядерных зон. Мелкими точками — третичные поля (зоны перекрытия)
Специфически человеческой особенностью являются существенные преобразования корковой организации, вызванные появлением второй сигнальной системы, связанной с трудовой деятельностью человека, речью, мышлением и сознанием. Преобразования, о которых здесь идет речь, сказываются на всех группах рассмотренных нами корковых полей. Особенно заметно они выражены в отношении вторичных и третичных полей коры.

Рис. 16. Схема выделения первичных, вторичных и третичных полей коры в сравнительно-анатомическом ряду млекопитающих (по
Г. И. Полякову): А — насекомоядные; Б — хищные и копытные; В — приматы и человек. Показано прогрессирующее выделение первичных (центральных) полей (обозначенных жирной штриховкой) наряду с ростом территорий периферических (вторичных) полей ядерных зон и образованием третичных полей — зон перекрытия (обозначены тонкой штриховкой)

Рис. 17. Соотношения роста различных территорий коры головного мозга человека в пост-натальном онтогенезе (сост. И. И. Глезером по данным Московского института мозга): ТРО — височно-теменно-затылочная область; Pi — нижнетеменная область; F — лобная область; ТМ — средняя височная подобласть; ТВ — базальные отделы височной области; Ос — затылочная область; TS — верхняя височная подобласть; Post — постцентральная область; lim — лимбическая область; ins — инсулярная область. Цифрами 0—10 обозначена степень увеличения соответственной области в постнатальном отногенезе
Только у человека в периферических отделах ядерных зон анализаторов и двигательной коры выделяются высоко специализированные участки, структурная и функциональная дифференцировка которых пошла в направлении анализа и синтеза раздражений, имеющих специальное значение в осуществлении различных форм речевой функции. Так, в заднем отделе периферического поля слуховой коры обособляется специальный участок для анализа и синтеза рецептивных элементов устной речи— фонем (так называемый «центр Вернике»), в периферических полях зрительной коры — участок, позволяющий реализовать анализ и синтез зрительных элементов рецептивной речи; в расположенных по соседству с кожно-кинестетической зоной участках нижних отделов теменной области, непосредственно примыкающих к чувствительным «центрам» руки, губ, языка, гортани, осуществляется анализ и синтез кожно-кине-сгетических рецепций, лежащих в основе артикуляции; в определенном отделе периферии двигательной зоны коры (нижние отделы лремоторной зоны) протекают нейродинамические процессы, связанные с преобразованием членораздельных звуков устной речи в сложные последовательные синтезы (так называемый «центр Брока»); в другом отделе премоторной зоны, примыкающем к двигательным «центрам» руки (в заднем отделе средней лобной извилины), локализуется программирование и осуществление сложных систем последовательных движений и двигательных навыков. Обо всем этом будет сказано в специальных разделах этой книги.

Рис. 18. Миэлогенетическая карта Фогтов. Крупными точками обозначены наиболее рано созревающие отделы коры. Мелкими точками — наиболее поздно созревающие отделы коры: а — наружная поверхность; б — внутренняя поверхность
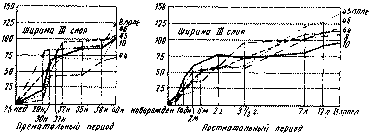
Рис. 19. Схемы роста ширины 3-го слоя в различных полях мозговой коры в онтогенезе (по Московскому институту мозга) Цифрами с правой стороны каждой схемы обозначены соответствующие поля; цифрами с левой стороны каждой схемы — степень роста ширины III слоя

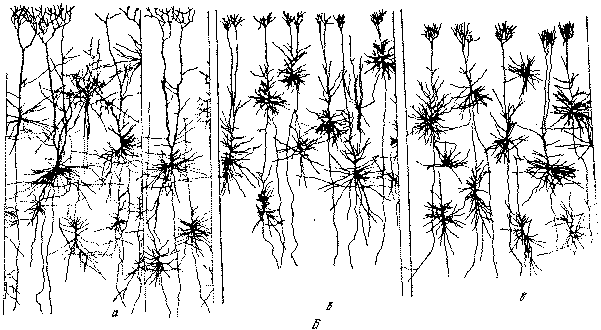
Рис. 20. Относительная тонкость нейронной структуры первичных, вторичных и третичных полей коры (по Г. И. Полякову): А — нейронное строение полей задних отделов полушария: а — первичное поле постцентральной области; б — вторичное поле постцентральной области; в — третичное поле (нижняя теменная область). Б — нейронное строение полей передних отделов полушария:а — первичное гигантопирамидное поле (двигательная область); б — вторичное поле (премоторная область); в — третичное поле (лобная область). На схеме видна нарастающая тонкость нейронной структуры при переходе от первичных полей к вторичным и от вторичных полей к третичным.
В связи с выделением в периферических полях ядерных зон специальных речевых отделов качественно совершенствуется и нейронная организация центральных полей ядерных зон, которая становится способной воспринимать и производить тончайшие чувствительные и двигательные дифференцировки элементов речевой функции. Этот процесс особенно наглядно выступает в центральном слуховом поле. Как показали исследования С. М. Блинкова (1955), это поле значительно возрастает у человека по сравнению с обезьяной как по своим абсолютным, так и по относительным размерам, обнаруживая при этом значительно дальше продвинувшуюся дифференцировку на ряд подполей. Эти прогрессивные изменения конструкции отражают фундаментальную роль устной речи как важнейшей основы деятельности всей системы словесной сигнализации.
Качественное преобразование, которое под влиянием формирования второй сигнальной системы испытывают зоны перекрытия анализаторов и лобной коры, сказывается в том, что все решительно сознательные психические процессы человека опосредуются через систему речевой сигнализации и протекают при доминирующем влиянии последней, о чем подробно говорится в последующих главах этой книги.
Наряду с дифференцировкой и качественной специализацией элементов всех слоев в филогенезе млекопитающих, и специально приматов, отмечается особенно значительное развитие слоя III коры, нейроны которого посылают, принимают и перерабатывают основную массу ассо-циационных импульсов коры. В ряду приматов ширина этого слоя прогрессивно нарастает от низших к высшим, все более отчетливо выявляется подразделение его на подслои. Сходный процесс имеет место и на протяжении всего периода онтогенетического развития коры.
Из представленных диаграмм (рис. 19) видно, что рост слоя III в ширину заметно обгоняет рост всех остальных слоев коры, составляя в соответствующих полях у взрослого человека в среднем около трети ширины всей коры.
Следует указать здесь еще на одну особенность нейронной организации, характеризующую все три группы корковых формаций и имеющую важное значение для понимания их физиологической роли в совокупной корковой деятельности. Дело в том, что характер нейронной организации в целом определенным образом изменяется при переходе от первичных полей к вторичным, а от них к третичным полям (рис. 20). Центральные корковые поля, через посредство которых замыкаются наиболее концентрированные потоки импульсов, связывающие кору наиболее непосредственным образом с органами чувств и органами движения на периферии, соответственно выделяются среди других полей относительной «грубостью» ' своей нейронной структуры, приспособленной для восприятия и обратной «отдачи» интенсивных возбуждений. Относительно наибольшей тонкостью нейронной структуры отличаются как раз третичные поля коры, что следует расценивать как морфологическое выражение особой тонкости и сложности функциональной дифференцировки этих полей, ответственных за наиболее высоко специализированные функциональные связи и взаимодействия между анализаторами. Вторичные поля занимают по всем особенностям переходное положение между первичными и третичными.
Описанный выше различный ход структурной дифференцировки полей задних отделов полушарий, имеющих отношение главным образом к организации разных форм ощущений и восприятий внешних раздражений, и полей передних отделов полушарий, имеющих отношение главным образом к организации действий, с большой отчетливостью выявляется также и в течение онтогенеза у человека. Было установлено, что уже начиная с ранних стадий эмбрионального развития коры процесс разделения на слои и формирование нейронов с их связями происходит в той и другой части полушарий отличным образом. В коре задней части полушария развитие слоев коры протекает по ясно выраженному типичному шестислойному типу, описанному Бродманом как гомотипическая кора, с обособлением концентрированного слоя IV как главного приемника афферентных импульсов из подкорковых отделов анализаторов. В коре всей передней части полушария, особенно в закладке двигательной зоны коры, слоистость внутри коры выступает в значительно более ослабленной форме, что соответствует более рассеянному расположению здесь (как и у взрослого человека) элементов, воспринимающих подкорковые импульсы. В закладке лобной коры обращает на себя внимание своеобразное расположение групп нейронов («волнистость»), обусловленное, очевидно, своеобразием развития межнейронных связей (рис. 21).

Рис 21 Различие структурной дифференциации коры передних и задних отделов в раннем онтогенезе у человека (по Г И Полякову) На схеме показаны два типа дифференцировки коры по поверхности полушарий в затылочной, височной и теменной областях хорошо выражена шестислойная структура коры; в лобной области — дифференцировка слоев коры протекает существенно отличным образом Буквенные обозначения E — эмбриональный тип коры, N — уплотненный II слой, и—волнистость; / — отсутствие дифференцировки внутри поперечника, // — разделение на верхний и нижний этажи, ///— выделение слоев в нижнем этаже, IV— выделение слоев также и в верхнем этаже
Дата добавления: 2015-10-16; просмотров: 168 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| В) Особенности структурной организации передних отделов коры | | | Д) Особенность структурной организации и связей медиобазальных отделов коры |