Читайте также:
|
Учение о мозговых механизмах произвольных движений претерпело за последнее время значительные изменения, связанные с общим пересмотром вопроса о строении двигательных процессов Исходной для этого пересмотра явилась рассмотренная выше рефлекторная концепция произвольных движений, которая заставила с новых позиций подойти к мозговым механизмам двигательного акта
Еще сравнительно недавно было принято считать, что корковый аппарат произвольных движений ограничен так называемой моторной зоной коры или 4-м полем Бродмана и соответствующими экотрапира-мидными (подкорковыми двигательными узлами Со времени исторической работы В. А. Беца (1874), открывшего гигантские пирамидные клетки в пятом слое этой зоны, установилось мнение, что постцентральная область является зоной чувствительных проекций, в то (Время как передняя центральная извилина представляет собой двигательный аппарат коры головного мозга, регулирующий произвольные движения и направляющий свои импульсы по эфферентному пирамидному тракту к передним рогам спинного мозга.
Рефлекторная концепция движений, сформулированная И. П. Павловым и его учениками (Н. И. Красногорским, 1911; С. Миллером и Ю. Конорским, 1936; Г. В. Скипиным, 1947, 1956, 1958) и разработанная Л. А. Орбели (1935), П. К. Анохиным (1935) и Н. А. Бернштей-ном (1947), заставила изменить эти представления.
Положение о том, что эфферентные двигательные импульсы являются лишь последним, заключительным этапом мозговой подготовки двигательного акта и что основную роль в ней играет система афферентных синтезов, формирующих эти двигательные импульсы, заменило понятие о «двигательных центрах» коры значительно более широким понятием «корковых отделов двигательного анализатора». Согласно этому новому представлению, в корковые отделы двигательного анализатора входит большой комплекс корковых зон, работа которых обеспечивает ту или иную сторону подготовки двигательного акта. Одни из них осуществляют организацию произвольных движений в системе внешних пространственных координат, другие анализируют импульсы, доходящие от мышечно-суставного аппарата, и, наконец, третьи обеспечивают то направляющее, регулирующее влияние речевых связей, которое, как мы уже указывали выше, является важнейшим составным элементом организации произвольного движения. Как это полагают некоторые авторы (Н. А. Бернштейн, 194 7; А. Р. Лурия, 1957), все эти области мозга должны быть включены в состав корковых отделав двигательного анализатора в широком смысле этого слова. Каждая из них, на чем мы еще остановимся ниже, вносит свой компонент в организацию двигательного акта, обеспечивая ту или иную стороны построения движения.
Однако наряду с широким понятием корковых отделов двигательного анализатора вполне законные основания имеет и более узкое представление о них, которое связано с выделением «коркового ядра» двигательного анализатора.
Особенность коркового ядра двигательного анализатора, отличающая его от соответствующих отделов других анализаторов, состоит в том, что к нему можно отнести не одну, а две совместно работающие зоны мозговой коры, которые обычно обозначаются как постцентральная и прецентральная области и которые вместе образуют единый функциональный аппарат — «сензомоторную область коры».
Такая особенность коркового аппарата двигательного анализатора имеет свои структурно-динамические и генетические основания. Структурно-динамические основания для такого выделения сензомоторной области, как коркового ядра двигательного анализатора, состоят в том, что самый двигательный а(кт представляет собой сложную функциональную систему. Для того чтобы осуществить произвольное движение, необходимо, чтобы зрительные, вестибулярные или слуховые импульсы были перешифрованы в известную систему кинестетических сигналов. Последняя составляет ту пространственную сетку, которая обеспечивает правильное направление эфферентных сигналов к соответствующим мышечным группам и которая вместе с тем динамически изменяет направление этих сигналов в зависимости от положения мышечно-суставного аппарата в пространстве. Без такой кинестетической основы никакое движение не может быть осуществлено. Это показывают известные опыты с деафферентацией конечности (произведенной на любом уровне афферентного аппарата), в результате которой исчезает четкость и диф-ференцированность двигательных импульсов. Эта кинестетическая основа движения и обеспечивается задними (постцентральными) отделами «коркового ядра» двигательного анализатора, перешифровывающими экстероцептивные сигналы в кинестетические и осуществляющими тот синтез импульсов в симультанные группы, о которых мы уже говорили выше (см. II, 3,в).
Кинестетический синтез является, однако, лишь одной стороной корковой организации произвольных движений. Каждое произвольное движение представляет собой не единичный пространственно организованный двигательный акт, но известную серию последовательных иннервации, протекающих во времени. Это можно наблюдать почти на любом движении и особенно отчетливо на каждом сложном двигательном навыке, который состоит из серии последовательно меняющихся иннервации, составляющих единую кинетическую мелодию. Эта кинетическая мелодия, как указывали многие авторы (Лешли, 1937; А. Р. Лурия, 1957 и др.), не может быть обеспечена тем же мозговым аппаратом, который ответствен за пространственную организацию двигательных импульсов, и по всем данным требует особого мозгового механизма, благодаря которому осуществляется денервация раз возникшего двигательного импульса и обобщение последовательных импульсов в единый, развертывающийся во времени двигательный стереотип. Есть много оснований для того, чтобы относить эту сторону эфферен тной интеграции двигательного акта за счет передних отделов «коркового ядра» двигательного анализатора и особенно за счет премоторной области (ом. ниже, II, 4, д).
На сложный характер коркового ядра двигательного анализатора, обеспечивающего как кинестетическую, так и «кинетическую» организацию движений, указывают генетические, анатомические и нейрофизиологические данные.
Как можно видеть из филогенетических исследований (Бродман, 1909, Г. И. Поляков, 1938—1948; Л. А. Кукуев, 1940; А. А. Хачатурян, 1949 и Др.), на ранних этапах филогенетического развития сензомоторная область составляла морфологически единую систему, которая лишь сравнительно поздно начала дифференцироваться на сензорную и моторную зоны.
Однако и после того, как это структурное единство сензомоторной области уступило свое место морфологической дифференцированности обеих областей, их функциональное единство продолжало существовать, проявляясь как в морфологических, так и в физиологических признаках.
Как было показано выше (см. I, 2, в), возбуждения, приходящие в постцентральную кору, переключаются на прецентральную двигательную кору либо через транскортикальные связи, либо же через вентро-латеральное ядро зрительного бугра, которое является подкорковым интегративным аппаратом двигательной системы. В связи с этим стоит и тот факт, что и в постцентральной и в прецентральной (двигательной) области мозговой коры имеются как афферентные, так и эфферентные комплексы волокон, что еще больше подчеркивает структурное и функциональное единство обеих частей «коркового ядра» двигательного анализатора. Если присоединить к этому важные данные Лассека (1954), показавшего, вопреки прежним представлениям, что лишь сравнительно небольшая часть волокон пирамидного пути начинается в гигантских бецевских клетках передней центральной извилины и что значительная их часть берет свое начало в постцентральных и премогорных отделах коры, единство указанных отделов сензомоторной коры выступит с еще большей отчетливостью.
Все эти данные заставляют по-новому отнестись к передней центральной извилине, которая долго рассматривалась как «двигательный центр» коры головного мозга. Гигантские пирамидные клетки Беца вне всяких сомнений являются аппаратами, генерирующими эфферентные двигательные импульсы, и входят в состав «передних рогов головного мозга» (Н. А. Бернштейн, 1947). Однако их нейрофизиологическая роль заключается не (Только в том, что они направляют возникшие в них импульсы к мышечной периферии. Она состоит еще и в том, что он«принимают на себя те многообразные динамические влияния, которые возникают в других афферентных отделах сензомоторной коры. Чем выше в эволюционном ряду стоит животное, тем более сложные системы афферентных импульсов 'принимаются бецевскими клетками. Вот почему, как это указал Бонин (1943), переход на более высокую эволюционную ступень (сопровождается увеличением бецевских клеток и сокращением их числа на один кубический миллиметр серого вещества, что приводит к (увеличению отношения всей массы серого вещества к массе тел бецевских клеток (см. таблицу).
Развитие структур коры передней извилины в филогенезе (по Бонину)
| Величина бецевских клеток (в микронах) | Число бецевских клеток на 1 мм3 серого вещества | Отношение массы серого вещества к массе тел бецевских клеток | |
| Низшие обезьяны Высшие обезьяны Человек | 3,7 - 6,1 | - |
Все рассмотренное выше дает полное основание считать, что корковым ядром двигательного анализатора является именно сензомотор-ная кора, морфологически дифференцированная, но выступающая как функциональное единство.
Это положение подтверждается и рядом физиологических наблюдений.
Как показали работы Мак-Кэллока (1943), выполненные нейро-нографическим методом, раздражение, приложенное к постцентральной коре, распространяется и на прецентральную нору, т. е. импульсы, возникающие от этого раздражения, можно проследить и в 4-м поле Бродмана. И обратно, раздражение, приложенное к прецентральной коре, распространяется на постцентральные ее отделы, и его эффект можно проследить в целом ряде полей, входящих в эту область (рис. 53).

Рис 53 Данные нейронографических опытов с раздражением постцентральных и прецентральных отделов коры (по Мак-Кэллоку). Схема показывает, как далеко распространяется возбуждение, вызванное раздражением стрихнином определенного участка коры головного мозга шимпанзе. Поля 6, 4, 3, 1, 5 и 7 относятся к сензомоторной коре. Заштри хованы так называемые депрессорные поля.
Вот почему, как это показали Пенфилд и Расмуссен (1950), раздражая «сензорные» отделы постцентральной области, можно получить не только чувствительный, но и двигательный эффект (рис. 54).
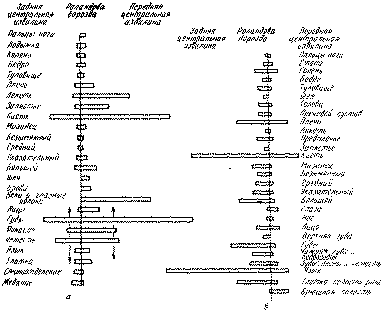
Рис 54 Двигательные и сензорные реакции при раздражении пре- и постцентральныхотделов коры (по Пенфилду и Расмуссену): α— представительство двигательных процессов в мозговой коре, б — представительство сензорных процессов в мозговой коре. Длина столбиков обозначает относительное число моторных и сензорных реакций, вызванных раздражением точек, расположенных спереди и сзади от Роландовой борозды.
Таким образам, мы имеем все основания рассматривать «двигательный центр» коры, в его узком значении, как (сложное образование, состоящее из двух совместно работающих областей (постцентральной а прецентральной), каждая из которых, как об этом будет сказано ниже, имеет свои специфические функции.
Топическая организация обеих зон, входящих в состав «коркового ядра» двигательного анализатора, имеет много общего с тем, что мы уже видели, рассматривая строение корковых отделов других анализаторов.
Центральное (или первичное) поле каждой из этих частей составляют проекционные участки коры головного мозга. Как известно, в проекционных участках постцентральной коры (поле 3-е Бродмана) кончаются волокна, идущие с периферии и несущие импульсы от одной из рецепторных зон, а в проекционном поле црецентральной области (поле 4-е Бродмана) начинаются волокна, идущие в составе пирамидного пути к двигательным нейронам шинного мозга. Характерно, что (проекции волокон в этих участках мозговой коры обладают отчетливым сома-тотопическим характером, так что нижние конечности представлены в верхних отделах контралатеральной сензомоторной коры, а верхние конечности и лицо — в средних и нижних отделах этой области. Пожалуй, наиболее существенная особенность (которую нельзя столь отчетливо видеть в корковых отделах других анализаторов) заключается в том, что (как это было сказано выше — см I, 2, б) проекция соответствующих отделов периферического сензомоторного аппарата представлена в указанных отделах сензомоторной коры не по геометрическому, а по функциональному принципу. Чем большее функциональное значение имеет тот или иной орган, чем более богаты его связи и больше его участие в системе произвольных движений, тем большую площадь занимает его проекция в коре головного мозга
На приведенном выше рис. 10 были представлены результаты опытов Пенфилда, в которых он, раздражая соответствующие участки коры сензомоторной области, получал чувствительные или двигательные ответы в связанных с ними участках тела Легко видеть, какое большое место занимает в корковых проекциях рука и органы речевого аппарата по сравнению с корковой проекцией туловища, бедра и ноги.
К первичным (проекционным) полям «коркового ядра» двигательного анализатора примыкают и вторичные поля 2, 1, 5, 7-е постцентральной и 6, 8-е поля Бродмана премоторной области По своему характеру они приближаются к вторичным полям «ядерной зоны» других анализаторов Как это будет показано ниже, они принимают ближайшее участие в процессах синтеза отдельных возбуждений в «группы» (постцентральная область) и в «ряды» (премоторная область), о которых мы уже неоднократно говорили.
Разбирая корковую организацию произвольных движений и их нарушения, мы выделяем две группы движений, которые представляют особый интерес для патологии высших психических функций а именно-движения руки, которые включены в различные формы произвольного действия, и движения орального аппарата, составляющие основу речи
Мы.последовательно разберем ту роль, которую играют,в организации этих движении «афферентные» (постцентральные) и «эфферентные» 1(премоторные) отделы коры, и рассмотрим те формы нарушений которые -возникают при поражении этих отделов мозга.
Весь этот аппарат, осуществляющий корковый анализ и синтез произвольных движений, надстраивается над аппаратом субкортикальных двигательных узлов, которые у человека не имеют самостоятельно-го значения обеспечивают лишь то, что некоторые авторы (Н. А. Бернштеин, 1947 и др.) называют «фоновыми компонентами двигательного акта». На симптоматике поражений этих образований мы специально останавливаться не будем.
1Термин "эфферентный" мы употребляем сдесь и нижев условном и далеко не точном смысле. Как будет видно из дальнейшего изложения, речь здесь идет также о своеобразной форме афферентации двигательного акта, но только эта афферентация носит совершенно особый характер.
Дата добавления: 2015-10-16; просмотров: 192 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| А) Исторический экскурс. Проблема | | | В) Нарушение афферентной основы произвольного движения и синдром «афферентной» (кинестетической) апраксии |