Читайте также:
|
(тесно связанно с гликолизом)
| СН2-ОН | СН2-ОН | |||
| Глицеролфосфаткиназа | │ | α-глицеролфосфатДГ | │ | |
| ГЛЦ |  
| СН2-ОН |  
| С=О |
| АТФ АДФ | │ | НАД НАДН2 | │ | |
| СН2-ОН | СН2-О-Ф | |||
| α-глицеролфосфат | дигидроксиацетон-фосфат (ДАФ) |
ДАФ далее подвергается анаэробному и аэробному гликолизу, в последнем случае включается в ЦТК, распадается до СО2 и Н2О или при потребности организма в углеводах, посредством реакций глюконеогенеза, пореобразуется в глюкозу.
β-окисление жирных кислот
происходит в печени. Впервые изучено в 1904 году Кнопом. В 1948-1949 гг. Кеннеди и Ленинджер установили, что β-окисление жирных кислот происходит в митохондриях. В 50-х годах Линеен описал ферменты окисления жирных кислот, поэтому процесс β-окисление жирных кислот называется циклом Кноопа-Линена.
ЭТАП
Предварительно жирные кислоты активируются в цитоплазме клетки (эндергонический процесс) т.е. процесс протекающий с повышением свободной энергии системы, требующий для своего осуществления приток энергии извне.

В результате образуется R-ацил-КоА, являющийся активной формой жирной кислоты.
ЭТАП
Мембрана митохондрий не проницаема для коэнзимная формы жирной кислоты, ее транспорт в митохондрии осуществляется с помощью карнитин а.

Реакция протекает при участии специфического цитоплазматического фермента карнитинацил-КоА-трансферазы. После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитинацил-КоА-трансферазы.
ЭТАП
Карнитин возвращается в цитоплазму клетки, а ацил-КоА подвергаются окислению в митохондриях
| О |  
|  
| О | ||||
| ║ | ║ | ||||||
| R - | С ~ SКоА | НО | - карнитин | R - | С ~ SКоА | ||
| HS - | КоА | R - | S ~ S - карнитин | HS - | КоА | ||
| цитоплазма | митохондрии | матрикс |
Стадия
1.дегидрирование

2. гидратация.

Стадия
3. дегидрирование
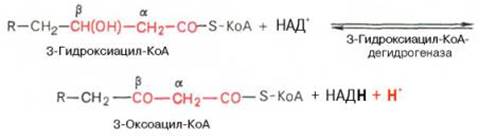 β-кетоацил КоА
β-кетоацил КоА
4.тиолазная реакция

Происходит расщепление β-кетоацил КоАна ацил-КоА и ацетил-КоА, укоротившийся на два углеродных атома.
Далее цикл повторяется до тех пор, пока не образуется бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА.
Ацетил-КоА → в ЦТК
НАДН2,ФАДН2 → в дыхательную цепь
Энергетический баланс β-окисления
(четное число атомов углерода)
При окислении жирной кислоты, содержащей n углеродныхатомов, происходит n /2–1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА происходит образование 2 молекул ацетил-КоА) и получится n /2 молекул ацетил-КоА. Следовательно, суммарное уравнение β-окисления пальмитиновой кислоты можно записать так:
Пальмитоил-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA ––> 8Ацетил-КоА + 7ФАДН2 + 7НАДН2.
При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают:
 7ФАДН2 х 2 = 14
7ФАДН2 х 2 = 14
7НАДН2 х 3 = 21 35 молекул АТФ (в дыхательную цепь)
8 ацетил-КоА х 12 = 96 молекулАТФ (в ЦТК)
ИТОГО: 35+96=131-1 (на образование активной формы жк) = 130 молекул АТФ.
Для ненасыщенной жк кол-во двойных связей х 2 = количество молекул АТФ.
| Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь АТФ характеризуется величиной 7,6 ккал/моль. Примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТФ, а оставшаяся часть, теряется в виде тепла. |
Дата добавления: 2015-08-17; просмотров: 199 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Внутриклеточный липолиз | | | Окисление жирных кислот |