|
Читайте также: |
ВСТУП
Рушійними факторами еволюції є нерозривно пов’язані між собою спадковість, мінливість і добір, що вперше було доведено Ч. Дарвіном (1859).
Наука про спадковість і мінливість живих організмів в онтогенетичному філогенетичному їх розвитку та методи управління цими процесами названа генетикою (від грецького genesis –походження). За відносно короткий період свого розвитку вона вийшла на передній край природознавства. Різноманітність життєвих форм і процесів може осмислена як єдине ціле лише в рамках генетики.
Для сучасної генетики характерним є, з одного боку, вплив на неї принципів і методів досліджень точних наук (математики, кібернетики, фізики, хімії), а також всезростаючий зв'язок її з іншими біологічними науками, з другого боку, сама генетика надзвичайно швидко диференціюється і окремі її розділи перетворюються в окремі науки. Так, за короткий період поряд із загальною генетикою, генетикою рослин і тварин виникли: цитогенетика, генетика мікроорганізмів, генетика вірусів, генетика хлоропластів, генетика мітохондрій, математична генетика, біохімічна генетика, космічна генетика, еволюційна генетика тощо.
Важливою рисою становлення сучасної генетики є те, що значний прогрес у вивченні спадковості і мінливості не заперечив, анітрохи не зменшив значення відкриттів, зроблених від її зародження до сьогодення. Навпаки, попередні відкриття ставали фундаментом для подальшого бурхливого розвитку, якого не переживала жодна наука за такий історично короткий період.
1. ІСТОРІЯ ВИВЧЕННЯ ГЕНЕТИЧНОГО
АПАРАТУ КЛІТИНИ
Численні прояви життя на Землі взаємопов’язані і мають спільні принципи організації, структури і функціонування.
Єдиною елементарною живою системою, якій властиві саморегуляція і самовідтворення, є клітина.Вона містить генетичну інформацію для наступного покоління.Клітина є матеріальною основою спадковості [3].
Вивчення клітини, а в подальшому і її генетичного апарату, пов’язане з відкриттям мікроскопа в 1665р. фізиком Робертом Гуком, який, розглядаючи тонкі зрізи пробки, відкрив дрібні порожнини і назвав їх клітинами. Але Р.Гук із своїх спостережень не зробив ніяких висновків [1].
В 1671 р. анатоми рослин Мальпігі і Грю одночасно і незалежно один від одного підтвердили відкриття Гука, показавши, що рослини складаються з тісно розташованих «пузирків» або «мішечків». Свою працю Мальпігі назвав «Огляд анатомії рослин», а Грю-«початком анатомії рослин» [1].
1680р. Антуаном Ван Левенгуком були відкриті різноманітні одноклітинні організми і тваринні клітини (еритроцити, сперматозоїди). Але перші вчені-цитологи надавали велике значення будові клітинної стінки, не звертаючи уваги на протопласт.І лише в 1831р. Робертом Броуном було відкрито ядро, який зазначив, що ядро є обов’язковим компонентом рослинної клітини і назвав його nucleus.Вивчення ядра він проводив на клітинах епідермісу.В 1837-1839рр. ядро описав Франц Мейен. Також в ці роки, німецькі вчені ботанік М.Шлейдон і зоолог Т.Шванн, вивчаючи будову тканин рослин і тварин, незалежно один від одного прийшли до висновку, що всі живі організми складаються з клітин.В 1855р. Р.Вирхов сформулював теорію, згідно з якою будь-яка клітина походить лише від клітини.Зоологи Шнейдр в 1873р. і Бючлі в 1874р. спостерігали картину поділу ядра в тваринних клітинах у червів, але не змогли узагальнити і оцінити побаченого явища.Це відкриття було зроблене російським вченим І.Д.Чистяковим 1874р.В 1875р. Е.Страсбургером також був детально описаний поділ ядра заплідненої яйцеклітини ялини [3].
В 1881р. італійським цитологом Є.Бальбіані були відкриті гігантські хромосоми.
У 1882р. німецький цитолог Ф.Флемінг побачив і описав зміни, які відбуваються в ядрі при кожному клітинному поділі.Після чого клітинне ядро стали вивчати особливо ретельно.
У 1886р. Е.Геккель висловив гіпотезу, що ядро клітини є носієм спадковості.
1888р. німецький вчений В.Вальдейтер вперше спостерігав хромосоми.
1901-1903рр. Гуго Де-Фріз голандський вчений висунув теорію мутації.
1909р. датський біолог В.Йоганин ввів у науковий обіг терміни «алель», «ген», «генотип», «фенотип».
1910р. Т.Г.Морган обґрунтував хромосомну теорію спадковості. Дослідження він проводив на плодовій мушці дрозофілі. Школа Моргана експериментально не тільки обґрунтувала хромосомну теорію, а й лінійне розташування генів у хромосомах та явище зчепленого успадкування. Також його учні А.Стертевант, К.Бріджисон, Г.Міллерс до середини 20-х років ХХст. сформулювали і перший варіант теорії гена-елементарного носія спадкової інформації. Проблема гена стала центральною генетики. Вона розробляється до цього часу.
1926р. німецький генетик Курт Штерн генетичними і цитологічними методами здійснив транс локацію фрагмента Y-хромосом на Х-хромосому у дрозофіли.Вперше було доведено локалізацію конкретного гена в хромосомі.
1927р. американський вчений Г.Меллер на прикладі дрозофіли експериментально довів можливість виникнення штучних мутацій під дією рентгенівських променів, при цьому він показав, що за такого впливу в сотні разів збільшується поява мутацій порівняно з природнім мутаційним процесом, що мутації є результатом випадкових молекулярних змін. Загалом, і представники школи Моргана, і інші вчені дали поштовх розвитку нового напрямку в генетиці – вчення про мутагенез.
На початку 30-х років ХХ-ст. В.В.Сахаров і М.Є.Лобашев отримали перші дані про виникнення спадкових змін під впливом деяких хімічних речовин.
А на рубежі 40-х років Дж.Бідл і Е.Татум започаткували основи генетики мікроорганізмів і біохімічної генетики. Вивчаючи механізми генетичного регулювання клітинного метаболізму, вони висловлювали припущення, що гени контролюють біосинтез ферментів.
1944р. О.Евері, К.Мак-Леод і М.Мак-Карті довели генетичну роль нуклеїнових кислот в експериментах з трансформації у мікроорганізмів. Вони виявили, що трансформуюим фактором було ДНК. Це відкриття символізувало початок нового етапу в розвитку генетики-зародження молекулярної генетики.
1946р. американські вчені Д.Ледеберг і Е.Тейтум довели наявність статевого процесу у бактерій. Процес перенесення генетичної інформації від однієї бактерії до іншої при контакті клітин дістав назву кон’югації.
1952р. М.Ціндер і Д.Ледерберг відкрили явище трансдукції-пернесення бактеріофагом генетичного матеріалу (ділянки ДНК) з однієї бактеріальної клітини в іншу, що спричиняє до змін спадкових властивостей клітин.
Епохальне відкриття зробили в 1953р. англійський біофізик і генетик Ф.Крик та американський біохімік Дж. Уотсон -спільно вони, розшифрували, створили модель структури ДНК (подвійну спіраль), що дозволило пояснити багато її властивостей та біологічні функції. Було встановлено, що гени є відрізками молекул ДНК.
В 1961р. американські вчені М.Ніренберг і Д.Маттеї відкрили генетичний код, довівши, що триплет нуклеотидів УУУ кодує амінокислоту феніл-аланін.
1961-1962рр. французькі мікробіологи-генетики Ф.Жакоб і Ж.Моно розробили схему регулювання біосинтезу білка і на її основі запропонували схему механізму генетичного контролю синтезу ферментів.
1968р. Г.Корана (США) синтезував активний штучний ген.
1970р. вчені Вісконского університету (США) відкрили фермент обернена транскриптаза, який здатний каталізувати синтез ДНК на матриксі РНК.
1972р. американський біохімік Пол Берг зі своїми співробітниками (Стенфордський університет)отримав перші рекомбінантні молекули ДНК двох різних вірусів, на яких досліджували структуру індивідуальних генів. Ці праці поклали початок народженню нової галузі- генної інженерії -системи експериментальних прийомів, які дають змогу створити штучні генетичні структури у вигляді рекомбінантних (гібридних) молекул ДНК.
1983р. Нобелівську премію було присуджено американському цитогенетику Барбарі Мак-Клінток за відкриття мігруючих елементів (мобільних генів) у генотипі кукурудзи [3].
Завдяки удосконаленню методів маніпуляції з генетичним матеріалом, розвивається принципово нові методи створення сортів рослин, порід тварин, генотерапії спадкових хвороб людини [8].
2. ПОНЯТТЯ ПРО ГЕНЕТИЧНИЙ АПАРАТ КЛІТИНИ
Клітина - це елементарна структурно-функціональна життєва одиниця. Вона визначає структуру та розвиток живих істот.
Ще в 1855р. Р.Вірхов висунув фундаментальне положення - «кожна клітина від клітини», в якому сконцентровано уявлення про само відтворення клітини [13].
Компоненти клітини, структурно-функціональна єдність яких забезпечує збереження, реалізацію та передачу спадкової інформації при розмноженні є генетичним апаратом клітини. Тривалий час вважали, що лише ядро є місцем локалізації генетичного матеріалу еукаріот. Лише на початку ХХ ст. Було знайдено ще такі органоїди цитоплазми як пластиди та мітохондрії, які також несуть певну кількість спадкової інформації. Генетичний матеріал організму-це сукупність носіїв його спадкової інформації-молекул ДНК і РНК [8].
Схематичне зображення організації генетичного апарату клітини предсталене на рис. 1.
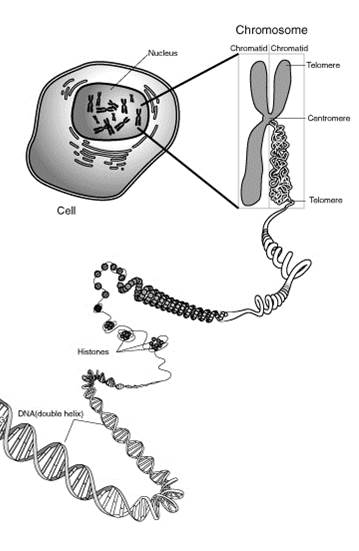 |
Будова ядра
Ядро –обовязкова частина будь-якої клітини як рослинної, так і тваринної. Від цитоплазми ядро відділене ядерною оболонкою, складається з двохмембран зовнішня ядерна мембрана зв’язана каналами ЕПС [1].
Ядерний сік, або каріоплазма, у виді неоструктурної маси оточує хромосоми і ядерця. Він кисліший за цитоплазму, що було визначено шляхом мікроін’єкції індикаторів.В ядерному сокові містяться ферменти, які беруть участь в синтезі нуклеїнових кислот, рибосом.
Ядерце - це постійна частина типового інтерфазного ядра, але відсутня під час поділу. За фізичними властивостями воно є найбільш щільною частиною ядра.В ньому міститься велика кількість РНК, кислих білків фосфопротеїнів. В ядерці відбувається синтез рибосом.
Хромосоми є основною функціональною частиною ядра [13].
2.2. Структура та морфологія хромосом
Хромосоми - це характерні структури генетичного матеріалу, який є обов’язковим компонентом ядра клітини. Вони мають особливу організацію, здатні до само відтворення і збереження своїх властивостей у кількох клітинних поколіннях. Кожна хромосома має певні морфологічні і генетичні особливості [6].
Хромосоми, знаходячись в ядрі, з давніх-давен цікавили дослідників як ймовірні носії закодованої спадкової інформації. Число, форма і розмір хромосом постійні у кожного індивідума і, загалом, у кожного виду. Прослідкувавши, як передають від покоління до покоління гени, контролюючи певні ознаки, можна упевнетись, що характер її передачі співпадає з характером передачі хромосом [1].
Хромосоми- (греч. chroma-колір, фарба, soma-тіло) -це окремі, добре помітні в оптичний мікроскоп структури, що формуються в профазі мітозу та мейозу завдяки спіралізації хроматину. Основу хромосоми становить одна безперервна дволанцюгова молекула ДНК. Довжина хромосом становить від 0, 2 до 20мкм [6].

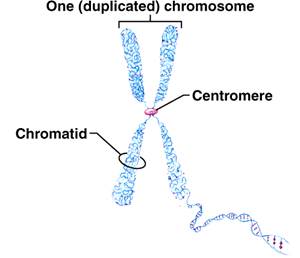 |
Рис. 2. Хромосома після реплікації ДНК. Рис. 3. Електронна мікрофотографія метафазної хромосоми
Тіло хромосоми розділяється первинною перетяжкою на два плеча. Первинна перетяжка (центромера, кінетохор) є світлішою, неспіралізованою ділянкою хромосоми. Центромера виконує функцію механічного центру хромосоми. Вона визначае орієнтацію хромосоми у мітозі і їх правильне розташування до полюсів. Якщо порушити тонким пучком ультрафіолетових промінів цю ділянку, то хромосоми втрачають рухомість в анафазі мітозу чи мейозу [10].
Зони хромосоми, що розміщені від центромери до половини довжини кожного плеча, називаються проксимальними. Зони другої половини плечей хромосоми, що знаходяться на дальшій віддалі від центромер, називаються дистальними.
Деякі хромосоми, крім первинної, мають ще й вторинну перетинку (нитку супутника), за допомогою якої утримується сегмент хромосоми – супутник. Деякі вторинні перетинки пов’язанні з утворенням ядерець – це ядерцеві організатори, в яких локалізуються гени, що відповідають за синтез р-РНК в ядерцях. Зустрічаються хромосоми, які на одному плечі мають подвійні (тандемні) супутники, що приєднуються двома вторинними перетяжками.
Кінцеві (термінальні) ділянки хромосоми називають – теломерами. Вони часто збагачені гетеро хроматином і забезпечують цілісність хромосом та перешкоджають злипанню їх кінців [4].
Структура хромосом починає проглядатися в профазі. На початку профази хромосоми мають вигляд тонких ниток. На цих нитках помітні темно забарвлені зернятка, або хромомери. Потім в профазі хромосоми потовщуються і метафазі являють собою вкорочені і потовщені утворення.При цьому вже в профазі спостерігається повздовжнє подвоєння хромосом – вони складаються з двох хроматид.
Хромосоми в інтерфазі складаються з ниток, які були названі хроматидами (від грецького – chroma – колір і eidos – подібний). Хроматиди утворені з нуклеїдних ниток хромонем, число яких у хроматид різноманітне від 2-1600 (у політенній хромосомі). Товщина кожної з них становить від 20-200 А. Хромонеми, в свою чергу, складаються з більш дрібних субодиниць – хромофібрил. Хромофібрили видно лише в електронному мікроскопі вони являють собою елементарні лінійні субодиниці і складаються з ДНК [6].
ДНК в хромосомах упаковано в нуклепротеїди, до складу яких входять також гістонові та негістонові білки (рис. 4).
2.3. Хімічний склад хромосом
Вивчення молекулярної будови хромосом призвело до висновку, що в основі їх лежать ДНК, РНК і білки [5].
На сьогодні, все з більшою ясністю стає відомо, що ДНК належить головна роль в передачі спадкових властивостей. Хімічна назва ДНК – дезоксирибонуклеїнова кислота; РНК – рибонуклеїнова кислота. Про будову ДНК
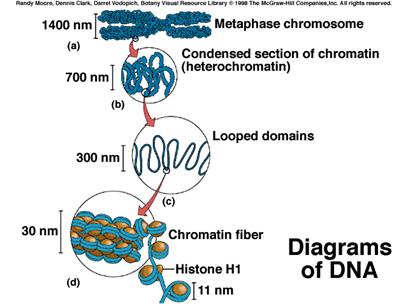 |
Рис. 4. Організація ДНК в метафазній хромосомі.
Крім нуклеїнових кислот, в склад хромосом входять білки – пістони; деякі ферментативні білки, наприклад, ДНК = полімераза, яка регулює реплікацію молекули ДНК в момент її синтезу. Також в хромосомах знайшли ліпіди і неорганічні комплекси: Са, Мg, Fe та інші. Вся ця конструкція хромосом на молекулярному рівні повністю ще не відома. Але є припущення, що віссю хромосом є ДНК в комплексі з білком, який отримав назву залишковий, тому що при видаленні гістонів конструкція хромосомів не руйнується. Одні дослідники припускають, що молекула ДНК в хромосомі нерозривна, інші ж допускають, що вона розривається ділянками іншої природи (білковими містками або неорганічними комплексами). Молекулярна, субмолекулярна і мікроскопічна структура хромосоми як єдиної цілої системи залишається до цих пір не ясною.
2.4. Еухроматин і гетерохроматин
Хромосоми по своїй осі не однорідні як хімічно так і фізично та генетично при фіксації основними фіксаторами різні ділянки, а й інколи цілі хромосоми дають різні реакції. Одні ділянки інтенсивно забарвлюються – їх називають гетерохроматиновими, інші – слабше забарвлюються, вони названі
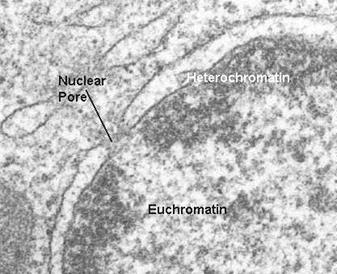 |
Рис. 5. Мікрофотографія еухроматинових та гетерохроматинових ділянок.
Гетерохроматинові і еухроматинові ділянки мають різні генетичні властивості. Перші майже не містять генів і спадково інертні, тоді як другі – складаються з генів і спадково активні. Штучне переміщення еухроматинових ділянок до інертних спричинює зміни прояву генів, які в них містяться. Гетерохроматинові ділянки розкидані по всій довжині хромосоми, але частіше розташовані поблизу центромери. В них частіше виникають розриви. Ніж у еухроматинових ділянках чи на межі з останніми [2,3].
Існують деякі види хромосом, які в основному складаються з гетерохроматину. До таких хромосом належать В- і У-хромосоми. Ці хромосоми, а також гетерохроматинові ділянки звичайних хромосом протягом всього клітинного циклу і особливо в інтерфазі можуть знаходитись у конденсованому – пікнотичному стані. У щитівок у такому стані можуть знаходитись цілі набори хромосом. Є припущення, що гетерохроматинові ділянки хромосом мають сильно спаралізований стан. Еухроматинові ділянки в інтерфазі деспіралізуються, що може вказувати на їх більш високу метаболітичну активність.
2.5. Політенні хромососми
Дуже цінною знахідкою для вчених цитологів та генетиків стала гігантська хромосома, яку вперше виявив італійський цитолог Є.Бальбіані в 1881р. у слинних залозах личинок хіраномуса. Пізніше така структура була виявлена в клітинах найпростіших, комах та синергідах деяких рослин, наприклад, у гороху. Найбільш типові гігантські хромосоми можна зустріти в слинних залозах мухи.
Утворюються такі хромосоми за рахунок того, що в клітинах алюмних залоз протягом періоду личинки не відбувається цитокінез, а реплікація хромосом здійснюється без їх розходження. Внаслідок такого ендомітозу хромосома набуває вигляду пучка хромонем, за що одержала назву політ енної.
Кожна гігантська хромосома може нараховувати до 800 хромонемних ниток, а завдяки соматичній кон’югації їх кількість збільшується вдвоє. При кон’югації гомологів хромонеми збігаються, утворюючи поперечні диски. Політенна хромосома в 100-200раз довша і в 1000раз товстіша від звичайних хромосом [5].
З відкриттям гігантських хромосом цитогенетика помітно збільшила можливості цитологічного контролю різноманітних генетичних процесів.
2.6. Каріотип
Сукупність набору хромосом клітини і, загалом, організму називають каріотипом. Каріотипування відіграє важливу роль у в ідентифікації хромосом. Для цього набір хромосом мета фазної клітини фотографують або змальовують за допомогою рисувального апарату. Після цього в результаті довгого ототожнення гомологічних хромосом складають каріограми організму.
Індивідуальна характеристика хромосом в каріотипі показує, що хромосомні набори індивідуальних видів різноманітні і відрізняються не лише по формі, а й по розміру. Аналіз каріотипів багатьох видів показав, що не існує в природі двох однакових каротипів, азначить і видів.
3. МОЛЕКУЛЯРНІ ОСНОВИ СПАДКОВОСТІ
3.1.Особливості будови ДНК
Розшифрування структури молекули ДНК, виконане американським вірусологом Дж. Уотсоном та англійським фізіологом Ф. Кріком в 1953р. допомогло їм створити молекулярну модель цього складного полімеру (рис. 6). Головні риси моделі були описані Уотсоном і Кріком, коли вони дійшли висновку, що ДНК – це високомолекулярний полімер, який складається з двох полі нуклеїнових ланцюгів [2].
 |
Мономерами кожного ланцюга є дезоксирибонуклеотиди. Розмір кожного нуклеотиду в поздовжньому вимірі молекули дорівнює 3.4 А0.
Ці ланцюги утворюють право закручені спіралі, кожний виток якої містить по 10 основ. Ланцюги полімеру закручені в спіраль один навколо другого і навколо загальної осі. Фосфатні групи цих ланцюгів містяться із зовнішньої сторони, а азотисті основи – всередині спіралі.
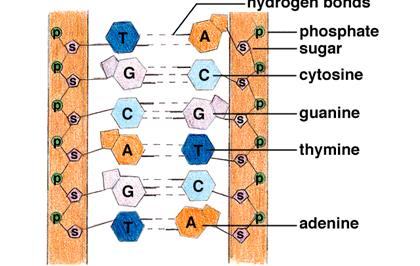
Довжина одного витка спіралі дорівнює 32 А0, а її діаметр – 20 А0. Ланцюги спіралі утримуються між собою за допомогою водневих зв’язків, які утворюються між парами аденін – тимінового (А - Т) та гуанін – цитозинового (Г- Ц) дизоксорибонуклеотидів. Пари А – Т та Г – Ц є специфічними в складі подвійної спіралі бо в нормі саме між гуаніном та цитозіном та між аденіном і тиміном у складі спіралі утворюються водневі зв’язки.
 |
Рис. 7. Нуклеотиди ДНК.
Всі типи нуклеотидів: аденіновий, гуаніновий, цитозиновий та тиміновий чергуються будови ланцюга в найрізноманітніших послідовностях. Послідовність головних валентностей нитки ДНК утворюються ефірними зв’язками між фосфорною кислотою і дезоксирибозоюю. Ці зв’язки утворюються атомами вуглецю молекулами цукру в 3’ та 5’ положенні.
Одна з важливих особливостей ДНК описується правилом еквівалентності, сутність якого полягає в тому, що в молекулах дезоксирибонуклеїнових кислот молярні відношення пуринів до піримідинів (або навпаки) дорівнюють одиниці (рис. 8). Ця закономірність відкрита Е.Чаргафом в 1950р. відіграла важливу роль у створенні дволанцюгової моделі молекули ДНК [3].
 |
Спираючись на створену модель, Уотсон і Крік дійшли висновку, що оскільки будова молекули полімеру монотонно повторюється цукрово-фосфатні послідовності, а мінливими виявляються тільки послідовності азотистих основ, що генетична інформація може бути закодованою лише за допомогою послідовностей азотистих основ, які входять до складу нуклеотидів у молекулах ДНК можуть спричиняти лише мутації. Отже, була виявлена специфічність взаємного розташування азотистих основ, яку почали розглядати як матеріальну основу генетичного коду.
Дата добавления: 2015-07-10; просмотров: 591 | Нарушение авторских прав