Вышеперечисленные структуры цитоплазмы объединены в единую группу по одной причине — элементарной составной их единицей является микротрубочка. Выявляются микротрубочки во всех типах клеток, за исключением бактерий. В структурах глаза, в частности в сетчатой оболочке, микротрубочки обнаруживаются в большом количестве. Основная функция микротрубочек — опорная, т. е. обеспечение определенной формы клетки и ее жесткости. По этой причине микротрубочки относят к структурам цитоскелета. Кроме того, они участвуют во внутриклеточном переносе метаболитов.

|
| 5 нм |
Микротрубочки имеют диаметр порядка 20—30 нм. Длина их различная. На поперечном срезе они имеют вид кольца (рис. 1.1.13). Каждая микротрубочка состоит из 13 протофила-ментов, расположенных вдоль длинной оси трубочки и скрученных по спирали одна над другой. Протофиламенты состоят из особого белка — тубулина. Сборка микротрубочки происходит из димеров тубулина (рис. 1.1.14). Синтез тубулинов происходит на мембранах грануляр-
Рис. 1.1.13. Схема структурной организации микротрубочки (по В. Л. Быкову, 1999):
а — мономеры тубулина, образующие протофиламенты; б — микротрубочка; s — пучок микротрубочек
Таблица 1.1.2. Морфо-функциональная организация митохондрий



 Структуры митохондрий
Структуры митохондрий
 Наружная мембрана Межмембранное пространство
Наружная мембрана Межмембранное пространство
Внутренняя мембрана
Субмитохондриальные частицы Матрикс
Состав
Около 20% всего белка митохондрий. Ферменты липидного обмена
Ферменты, использующие АТФ для фосфорилирования других нуклеоти-дов
Ферменты дыхательной цепи, цито-хромы. Сукцинатдегидрогеназа. Трансбелки.
АТФ-синтетаза
Ферменты (кроме сукцинатдегидро-геназы). ДНК, РНК, рибосомы, ферменты, участвующие в экспрессии генома митохондрий
Функция
Транспорт. Превращение липидов в промежуточные метаболиты
Создание электрохимического протонного градиента. Перенос метаболитов в матрикс и из него
Синтез и гидролиз АТФ
Цикл лимонной кислоты, превращение пирувата, аминокислот и жирных кислот в ацетил-коэнзим А. Репликация, транскрипция, трансляция
Клетка


|
мирующих сети. Микротрубочки могут образовывать пучки, в которых они связаны тонкими поперечными мостиками (в отростках нейронов, в составе митотического веретена и др.). Нередко микротрубочки частично сливаются, формируя пары (в аксонеме ресничек и жгутиков) или триплеты (в базальном тельце и цент-риоли).
| 12 |
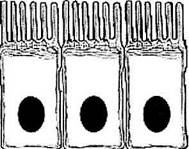
 Микротрубочки являются составной частью и другого органоида—реснички (рис. 1.1.14, 1.1.15). Реснички располагаются на апикальной поверхности многих клеток, в основном эпителиальных, выстилающих влажные поверхности тканей. В клетке может быть одна или несколько сотен ресничек. Обычно ресничка имеет длину порядка 15 мкм, а диаметр — 0,2 мкм. В основании реснички располагается электрон-ноплотное образование, называемое базальным тельцем. Базальное тельце цилиндрическое и состоит из девяти пучков параллельных друг другу микротрубочек, по три в каждом пучке. Такой пучок, состоящий из трех микротрубочек, называется триплетом. Девять триплетов удерживаются фибриллярным материалом, образуя стенку цилиндра.
Микротрубочки являются составной частью и другого органоида—реснички (рис. 1.1.14, 1.1.15). Реснички располагаются на апикальной поверхности многих клеток, в основном эпителиальных, выстилающих влажные поверхности тканей. В клетке может быть одна или несколько сотен ресничек. Обычно ресничка имеет длину порядка 15 мкм, а диаметр — 0,2 мкм. В основании реснички располагается электрон-ноплотное образование, называемое базальным тельцем. Базальное тельце цилиндрическое и состоит из девяти пучков параллельных друг другу микротрубочек, по три в каждом пучке. Такой пучок, состоящий из трех микротрубочек, называется триплетом. Девять триплетов удерживаются фибриллярным материалом, образуя стенку цилиндра.
6 2
| ной эндоплазматической сети, а сборка в спирали — в клеточном центре. При этом поддерживается постоянное равновесие между сформированной микротрубочкой и растворенными в цитоплазме димеров тубулина, способных к самосборке. Эта закономерность не распространяется на постоянные органоиды клеток, состоящие из микротрубочек, — реснички, цент-риоли, базальные тельца. Нарушают процесс самосборки некоторые вещества, в частности колхицин и винбластин. Микротрубочки являются структурным компонентом веретена при делении клетки. Микротрубочки формируют в цитоплазме различные структурные системы. Они могут быть распределены в виде отдельных элементов, разбросанных по всей цитоплазме и фор- |
Рис. 1.1.14. Ультраструктурные особенности ресничек: а — продольный срез; б — поперечный срез
Рис. 1.1.15. Схематическое изображение организации реснички (по В. J1. Быкову, 1999):
а —продольный срез; б — поперечный срез (/ — базальное тельце; 2 — центр организации микротрубочек; 3 — базальный корешок; 4 — плазмолемма; 5 — микротрубочка А; 6 — микротрубочка В; 7 — периферические микротрубочки; 8 — центральные микротрубочки; 9 — центральная оболочка; 10 — динеино-вые ручки; // — радиальные спицы; 12 — нексиновые мостики)
Базальное тельце является организатором реснички. После образования базального тельца оно мигрирует к апикальной поверхности клетки. Из дистального конца базального тельца растут микротрубочки, составляющие стержень реснички (аксонема). Этот стержень, окруженный цитоплазматической мембраной, и вы-
Глава 1. КЛЕТКА И ТКАНИ
 стоит над поверхностью клетки. В центре аксо-немы образуются две одиночные микротрубочки, называемые центральной парой, или синг-летами.
стоит над поверхностью клетки. В центре аксо-немы образуются две одиночные микротрубочки, называемые центральной парой, или синг-летами.
Образованные реснички синхронно совершают движения, способствуя продвижению по эпителиальной поверхности слизистой оболочки секрета. Реснички эндотелия роговой оболочки обеспечивают перемещение камерной влаги в определенном направлении и с определенной скоростью, что имеет немаловажное значение в метаболизме структур глаза. Напоминающая ресничку структура с базальным тельцем обнаруживается и в фоторецепторных клетках.
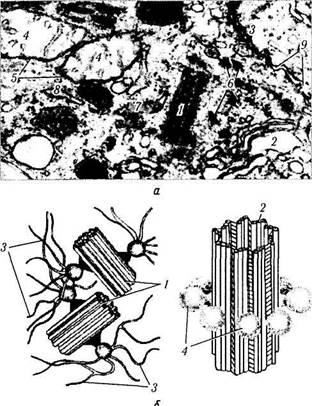
Следующей структурой, состоящей из микротрубочек, является клеточный центр, образованный двумя полыми цилиндрическими структурами. Длина клеточного центра равна 0,3—0,5 мкм, а диаметр — 0,15—0,2 мкм. Каждая из этих структур называется центриолью. Располагаются они вблизи друг друга во взаимно перпендикулярных плоскостях недалеко от аппарата Гольджи (рис. 1.1.16). Часть цитоплазмы, где они лежат, называется центросомой.
И
Рис. 1.1.16. Клеточный центр и структурная организация центриоли:
а —ультраструктурные особенности клеточного центра (/— центриоль; 2 — комплекс Гольджи; 3 — десмосома; 4 — митохондрия; 5 — мембрана митохондрии; 6 — микротрубочки; 7 — шероховатый эндоплазматический ретикулум; 8 — гладкий эндо-плазматический ретикулум; 9 — межклеточное пространство) б — схема организации клеточного центра и центриоли (/ — центриоли; 2 — триплеты микротрубочек; 3 — микротрубочки; 4 — сателлиты)
Ультраструктурная их организация практически неотличима от строения базального тельца. Каждая центриоль состоит из 9 триплетов частично слившихся микротрубочек, связанных поперечными мостиками. Каждый триплет связан со сферическими тельцами (сателлитами). Расходящиеся от них микротрубочки образуют центросферу.
В неделящейся клетке выявляется одна пара центриолей (диплосома). Перед делением (S-фа-за) происходит дупликация центриолей пары, причем под прямым углом к каждой зрелой центриоли формируется новая (дочерняя), незрелая процентриоль. Пары центриолей затем расходятся к полюсам клетки. Во время митоза они служат центрами образования микротрубочек ахроматического веретена деления. Таким образом, основной функцией центриолей является участие в митотическом делении клетки.
Филаменты. В цитоплазме большинства клеток обнаруживается множество волокнистых структур (филаментов) (рис. 1.1.17, 1.1.18). Различают три типа филаментов (микрофила-менты, миозиновые филаменты и промежуточные филаменты).
Первый тип филаментов — это так называемые микрофиламенты. Диаметр их 5—б нм. В основном они состоят из белка актина. С актином связываются еще два типа белка, а именно тропомиозин и миозин. В результате этого процесса формируется актино-миозино-вый комплекс. При этом актин и миозин становятся способными смещаться в этом комплексе продольно относительно друг друга. Если концы комплекса скреплены с какими-либо другими внутриклеточными структурами, последние сближаются. Этот процесс лежит в основе перемещения внутри цитоплазмы органоидов, транспортных пузырьков и других структур. На этом основано и мышечное сокращение.
Микрофиламентов особенно много в поверхностных областях цитоплазмы (поверхностный комплекс). Тем самым они способствуют поступлению веществ в цитоплазму (пиноцитоз), обладая возможностью изменять конфигурацию плазмолеммы.
Актиновые филаменты прикрепляются к трансмембранным белкам в особых участках плазмолеммы, называемых адгезионными соединениями, или фокальными контактами, которые связывают клетки друг с другом или клетки с компонентами межклеточного вещества.
Второй тип филаментов называют миозино-выми филаментами, поскольку они состоят из белка миозина. Этот тип филаментов тесно связан с актиновыми микрофиламентами в мышечных клетках. Эти филаменты толще (диаметр равняется 10 нм).
Название третьего типа филаментов — промежуточные. Их диаметр колеблется от 7 до 10 нм. Промежуточные филаменты наиболее часто встречаются в нервных и глиальных клет-

|

|
|
Клетка

 25 мкм
25 мкм

25 нм
Рис. 1.1.17. Особенности распределения структурных элементов цитоскелета (верхняя часть рисунка) и их
молекулярная организация (нижняя часть рисунка):
а — промежуточные филаменты; б — микротрубочки; в — актиновые филаменты
 |
| Необходимо отметить и то, что, несмотря на сходное строение, промежуточные филаменты отличаются в клетках различных тканей своим химическим составом (табл. 1.1.З.). Таблица 1.1.3. Распределение промежуточных филаментов различных классов в клетках и тканях человека |
| Химические особенности белков промежуточных филаментов легко демонстрируются иммуноцитохимическими методами, поскольку каждый белок является антигеном. Идентификация классов промежуточных филаментов имеет большое диагностическое значение при установлении тканевой принадлежности опухоли. Наибольшее диагностическое значение |

Рис. 1.1.18. Продольный срез отростка глиальной клетки сетчатки. Ультраструктурные особенности внутри-цитоплазматических филаментов
ках (в частности, в сетчатой оболочке, зрительном нерве). Эти филаменты в клетке образуют трехмерные сети. Входят они также в состав десмосом и полудесмосом эпителиальных клеток. К основным функциям промежуточных филаментов в настоящее время относят опорную функцию, обеспечение равномерного распределения сил деформации между клетками ткани (препятствует повреждению отдельных клеток), участие в образовании рогового вещества в эпителии кожи, поддержание формы отростков нервных клеток и фиксация трансмембранных белков. Кроме того, эти филаменты обеспечивают удержание миофибрилл в мышечной ткани и прикрепление их к плазмо-лемме (обеспечение сократительной функции мышц).
Классы промежуточных филаментов
 (Цито-)кератиновые (тонофиламенты)
(Цито-)кератиновые (тонофиламенты)
Десминовые
Виментиновые
Нейрофиламенты
Глиальные (содержат глиальный фибриллярный кислый белок)
Ламины (образуют ка-риоскелет)
Типы клеток и тканей
Эпителиальные
Мышечные ткани — гладкие (кроме миоцитов сосудов) и поперечнополосатые
Различные клетки мезенхим-ного происхождения: фибро-бласты, макрофаги, остеобласты, хондробласты, эндотелий и гладкие миоциты сосудов
Нейроны
Глиальные клетки (астроциты, олигодендроглиоциты)
Все типы клеток
Глава I. КЛЕТКА И ТКАНИ

 имеет выявление цитокератинов, десмина и глиального фибриллярного кислого белка, которые служат маркерами опухолей эпителиального, мышечного и глиального происхождения. Менее отчетливые результаты дает обнаружение виментина.
имеет выявление цитокератинов, десмина и глиального фибриллярного кислого белка, которые служат маркерами опухолей эпителиального, мышечного и глиального происхождения. Менее отчетливые результаты дает обнаружение виментина.
Микроворсинки. Некоторые (эпителиальные) клетки на своей апикальной поверхности содержат многочисленные цитоплазматические выросты, значительно увеличивающие площадь контакта содержимого цитоплазмы с окружающей средой, что способствуют увеличению интенсивности всасывания питательных веществ. Называются эти органоиды микроворсинками (рис. 1.1.5, 1.1.19). Естественно, что чаще подобные образования обнаруживаются в эпителиальных клетках, особенно клетках слизистой желудочно-кишечного тракта. Пигментный эпителий сетчатой оболочки также обладает подобными образованиями, распространяющимися между наружными члениками палочек и колбочек фоторецепторных клеток сетчатки. Многочисленны они и на апикальной поверхности эндотелиальных клеток роговой оболочки.
Микроворсинки имеют диаметр порядка 0,1 мкм. Длина их может быть самой различной.
В центральной части микроворсинки, представляющей собой выпячивание цитоплазмы, располагается порядка 40 микрофиламентов (диаметр 6 нм). В апикальной части микроворсинки пучок микрофиламентов закреплен в аморфном веществе. Его жесткость обуслов-
Рис. 1.1.19. Схема ультраструктурной организации микроворсинки:
/ — актиновые микрофиламенты; 2 — аморфное вещество; 3 — фимбрин и виллин (белки, образующие поперечные сшивки в пучке актиновых микрофиламентов); 4 — молекулы минимиозина (прикрепляющие пучок актиновых микрофиламентов к плазмолемме микроворсинки); 5 — терминальная сеть актиновых микрофиламентов; 6 — спектриновые мостики (прикрепляют терминальную сеть к плазмолемме); 7 — миозиновые филаменты; 8 — промежуточные филаменты; 9 — гликокаликс
лена наличием поперечных сшивок из белков фимбрина и виллина. Изнутри пучок прикреплен к плазмолемме микроворсинки белковыми мостиками из минимиозина.
Непосредственно под ворсинкой в цитоплазме обнаруживается скопление миозиновых фи-ламентов. Предполагается, что взаимодействие этих двух типов филаментов способствует изменению конфигурации микроворсинок, что еще более усиливает всасывательную функцию клеток.
Стереоцилии представляют собой видоизмененные длинные микроворсинки.
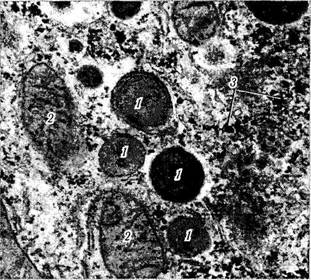
Лизосомы (цитосомы). Уже давно в большинстве клеток были обнаружены мембранные органоиды в виде «темных телец» различного размера (0,4—0,5 мкм) (рис. 1.1.20). Основным отличием этих образований было то, что они содержали целый набор (около 50) гидролаз. Поскольку гидролазы осуществляют лизис, эти органоиды и были названы «лизосомы» (переваривающие тельца).
Рис. 1.1.20. Лизосомы:
а — высокая степень насыщения цитоплазмы лизосомами, вблизи которых видны бобовидной формы митохондрии; б — ультраструктурные особенности вторичной лизосомы (/ — электронно-плотные включения в лизосому; 2 — мембрана лизосомы; 3 —митохондрии; 4 — гладкий эндоплазматический ретикулум)
Клетка

|
| Пиноцитоз-ные пузырь- Мультивезику- / ки Аутофаго- лярное тельце / сомы /ffcv\\ |
| Фагоцитоз |
| Первичная лизосома |
| ш Фагоцитоз |
| Комплекс Гольджи |
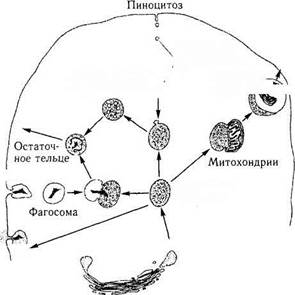
Лизосомы могут быть идентифицированы в срезах только при помощи гистохимических методов исследования, выявляющих активность двух или нескольких ферментов. Синтезируются ферменты на рибосомах гранулярной эндо-плазматической сети, переносятся транспортными пузырьками в аппарат Гольджи, где и модифицируются. От зрелой поверхности аппарата Гольджи отпочковываются первичные лизосомы (рис. 1.1.21).
Рис. 1.1.21. Схема, иллюстрирующая фагоцитоз, пино-
цитоз и образование первичных лизосом из комплекса
Гольджи (по А. Хэм, Д. Кормак, 1982)
Наличие гидролитических ферментов предопределяет и основную функцию лизосом. Они расщепляют вещества различного строения как внутри-, так и внеклеточного происхождения. При переваривании инородных веществ внутри клетки ферменты не выходят за пределы мембран лизосом. Лишь при патологических условиях наступает разрушение мембраны лизосомы и ферменты высвобождаются в цитоплазму. Действие ферментов приводит к лизису содержимого цитоплазмы, гибели клетки. Этот процесс называется аутолизом.
Лизосомы играют большую роль в поддержании нормального метаболизма клетки, защите организма от бактерий, токсинов.
Термин «вторичные лизосомы» используют в тех случаях, когда в цитоплазме клетки обнаруживаются так называемые пищеварительные вакуоли. Такие вакуоли возникают при поглощении клеткой чужеродных веществ (фагоцитоз). При этом первоначально происходит поглощение, а затем и переваривание их. В результате этого образуется «остаточное тельце», которое выталкивается из клеток путем экзоцитоза.
Необходимо остановиться и на роли лизосом в физиологической регенерации внутриклеточных структур. Этот процесс происходит следующим образом. Вблизи поврежденных или требующих замены участков цитоплазмы образуется полулунная двойная мембрана, которая растет и окружает со всех сторон поврежденные зоны. Затем эта структура сливается с ли-зосомами. В такой аутофагосоме совершается лизис структур органеллы. Таким образом, аутофагия представляет собой один из механизмов обновления внутриклеточных структур — внутриклеточной физиологической регенерации.
В глазном яблоке большее количество лизосом выявляется в пигментном эпителии сетчатой оболочки. Именно здесь они принимают участие в физиологической регенерации наружных члеников палочек и колбочек (см. Сетчатка).
Нарушение функции лизосом приводит к развитию ряда заболеваний, называемых лизо-сомными болезнями. Дефицит (чаще врожденный) ряда гидролитических ферментов лизосом приводит к накоплению в клетках непереваренных продуктов обмена (чаще всего гликогена, гликолипидов, гликозаминогликанов), нарушающих функцию клетки (болезни накопления). Нередко при таких заболеваниях поражается центральная нервная система и зрительный анализатор.
| Рис. 1.1.22. Ультраструктурные особенности пероксисом: / — пероксисомы; 2 — митохондрии; 3 — зерна гликогена |
Пероксисомы. Пероксисомы представляют собой мембранные пузырьки диаметром от 0,05 до 1,5 мкм (рис. 1.1.22). Они отщепляются от цистерн транс-полюса аппарата Гольджи. Различают две формы пероксисом. Мелкие пероксисомы (0,15—0,25 мкм) обнаруживаются во всех клетках млекопитающих. Крупные (более
Глава 1. КЛЕТКА И ТКАНИ
 0,25 мкм) присутствуют лишь в некоторых тканях (почки, печень). В них обнаруживается кристалловидная сердцевина, в которой находятся ферменты в концентрированной форме.
0,25 мкм) присутствуют лишь в некоторых тканях (почки, печень). В них обнаруживается кристалловидная сердцевина, в которой находятся ферменты в концентрированной форме.
Пероксисомы содержат около 15 ферментов (пероксидаза, каталаза и оксидаза D-аминокис-лот). Пероксидаза участвует в обмене перекис-ных соединений, часть которых токсична для клетки (перекись водорода). Пероксисомы участвуют в нейтрализации многих токсических соединений, в обмене липидов, холестерина, пуринов.
В настоящее время открыт новый класс наследственных заболеваний человека, насчитывающий не менее 12 нозологических форм— пероксисомные болезни. Развитие этих заболеваний связано с дефектом активности перокси-сом. При этих заболеваниях поражается центральная нервная система, и заболевание приводит к смерти в раннем возрасте.
Меланосомы. Меланосомы представляют собой органоид, основной функцией которого является синтез пигмента — меланина. Эти органоиды обнаруживаются в клетках меланоци-тарной системы, к которым относятся стро-мальные меланоциты кожи и некоторых слизистых (бульбарная конъюнктива, слизистая толстого кишечника), меланоциты увеального тракта глаза человека (стромы радужной оболочки, ресничного тела, хориоидеи). Вышеперечисленные клетки происходят из клеток нервного гребня путем их миграции на ранних этапах эмбриогенеза.
Меланосомы обнаруживаются также в клетках нейроэпителиального происхождения (ней-ромеланин). К таковым относятся клетки пигментного эпителия радужки, ресничного тела и сетчатки. Меланосомы обнаруживаются также в некоторых нейронах головного мозга — нейроны черной субстанции.
Меланин, продуцируемый меланосомами, представляет собой темно-коричневый пигмент (рис. 1.1.23), интенсивно поглощающий световую энергию, особенно коротковолоновой части спектра (ультрафиолетовую энергию, 290—320 нм). Меланин обладает способностью «гасить» свободные радикалы. Благодаря этим способностям меланин предохраняет ткани от повреждающего действия ультрафиолетовой энергии.
Формирование меланина происходит в ме-ланосомах меланоцитов под действием фермента тирозиназы, который преобразует аминокислоту тирозин в дигидрооксифенилала-нин (ДОФА) с последующим превращением в ДОФА-квинон. Затем происходит полимеризация ДОФА-квинона с формированием зерен меланина (рис. 1.1.23, 1.1.24).
Темно-коричневый меланин называют эуме-ланином, а меланин красноватого цвета — фео-меланином. Эти два типа меланина различаются и химическим составом. Красноватый
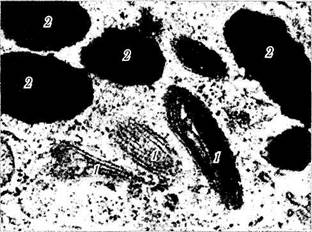
Рис. 1.1.23. Различные стадии формирования мелано-сом (трансмиссионная электронная микроскопия):
1 — премеланосомы; 2 — меланосомы

Рис. 1.1.24. Схема, изображающая стадии формирования меланосом:
х — тирозиназа; стадии I, II, III, IV
пигмент отличается включением в его состав серосодержащей аминокислоты с образованием 5-цистенил-ДОФА.
Как указывалось выше, активность тирозиназы проявляется в меланосомах. Последние представляют собой овальные или округлые тельца, диффузно распределенные в цитоплазме меланоцитов или клеток пигментного эпителия радужки, ресничного тела, сетчатки. Меланосомы образуются в результате биосинтетической деятельности гладкого и шероховатого эндоплазматического ретикулума, а также комплекса Гольджи [33]. Выделено четыре стадии меланизации меланосом. На первой стадии ме-ланосома представляет собой пузырек, содер-
Клетка

|
| Кератиноциты |
жащий тирозиназу, но не меланин (рис. 1.1.25). На второй стадии меланосома превращается в овальную структуру, выполненную нежным фи-ламентозным материалом. На третьей стадии этот филаментозный материал начинает мела-низироваться. В четвертой стадии наступает полная меланизация.
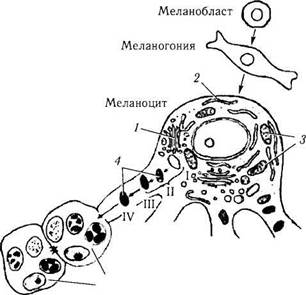
Рис. 1.1.25. Схема, иллюстрирующая развитие мелано-цитов, их дифференциацию и взаимодействие с эпителиальными клетками эпидермиса (по Jimbow et ai, 1976):
1 — комплекс Гольджи; 2 — эндоплазматический ретикулум; 3 — митохондрии; 4 — меланизация меланосом (стадии I, II, III и IV)
Количество меланосом, степень их мелани-зации, количество меланоцитов и определяют степень пигментации кожи и структур глаза. Размер меланосом частично находится под генетическим контролем. Так, размер меланосом у негров колеблется от 1,0—1,3 мкм, а у белых— 0,6—0,7 мкм [31]. Различия обнаруживаются в стадийности процесса. У белых индивидуумов в большем количестве обнаруживаются меланомы в стадиях / и //, а у негров — в стадии IV.
В коже меланин передается клеткам эпидермиса посредством фагоцитоза эпителиальной клеткой зерен меланина, распространяющихся к ним по цитоплазматическим отросткам мела-ноцита [36]. В кератиноцитах зерна меланина по мере дифференциации клеток смещаются к поверхностным слоям. В эпителиальных клетках меланоциты формируют агрегаты, окруженные мембраной. Эти структуры напоминают вторичные лизосомы. В них происходит частичная деградация меланина. Оставшийся меланин удаляется в результате слущивания поверхностных клеток эпидермиса.
Выведение зерен меланина в увеальных ме-ланоцитах и пигментных клетках сетчатки про-
исходит путем фагоцитоза выделившихся в результате распада меланоцитов зерен меланина макрофагами (меланофаги). Последние мигрируют по направлению кровеносных сосудов, проникают в их просвет и высвобождают меланин. В случаях нарушения этих процессов происходит накопление внеклеточно расположенного меланина. При высвобождении меланина в камерную влагу (дегенерация пигментного эпителия радужки, ресничного тела, посттравматические изменения) зерна меланина выводятся через дренажную систему глаза. В тех случаях, когда меланина большое количество, возможна блокада трабекулярной сети, в результате чего развивается так называемая «пигментная глаукома».
В заключение необходимо отметить, что синтезирующие меланин клетки — меланоциты — являются источником одних из наиболее злокачественных опухолей — меланом (кожи, слизистых, конъюнктивы, увеального тракта глаза). Одним из наиболее важных диагностических признаков этого заболевания является выявление при микроскопическом исследовании меланосом (иногда при помощи импрегнации серебром методом Фонтана) или активности ти-розиназы (в случаях отсутствия меланизации меланосом).
1.1.2. Внутрицитоплазматические включения
Гликоген. В цитоплазме многих клеток обнаруживаются неправильной формы частицы диаметром 20—30 нм. Различают два типа гликогена. Первый тип характеризуется скоплением изолированных частиц, равномерно или неравномерно распределенных в цитоплазме (бета-частицы). Помимо бета-частиц можно обнаружить и альфа-частицы. Отличия сводятся к тому, что в альфа-частицах отдельные гранулы собираются в розетки. Наиболее часто гликоген обнаруживается в клетках печени и мышечной ткани.
Липиды. В норме в цитоплазме клеток обнаруживаются капельки липидов. Депозиты липи-дов имеют различный вид. Часть капелек гомогенного вида, в то время как другие обладают пластинчатой структурой. Отличаются они и плотностью. Большинство свободных липидов исчезает в процессе гистологической обработки материала, поскольку вымываются спиртами и ксилолом.
Пигментные гранулы. В тканях глаза можно обнаружить два типа пигментных гранул. Это меланин и липофусцин.
Меланин широко представлен в тканях глаза, выполняя довольно важные функции в фоторецепции. Меланиновые гранулы обнаруживаются как в нейроэпителиальных производных, таких как пигментный эпителий сетчатки, радужки, ресничного тела, так и в стромальных
Глава 1. КЛЕТКА И ТКАНИ

|

|
меланоцитах увеального тракта (рис. 1.1.23). Пигментные гранулы перечисленных двух оболочек довольно трудно различать при световой микроскопии. Гранулы, выявляемые в нейро-эпителии, несколько больше в размерах и темнее, чем обнаруживаемые в увеальном тракте. Второй тип гранул — липофусцин — может быть спутан с меланином, поскольку он имеет светло-коричневый цвет и по размерам приближается к зернам меланина (рис. 1.1.26). Гранулы липофусцина имеют более светлый цвет. Липофусцин чаще обнаруживается в пожилом и старческом возрасте, особенно в клетках миокарда и нейронах. Довольно много появляется его с возрастом в клетках пигментного эпителия сетчатки.
Рис. 1.1.26. Зерна липофусцина (стрелки) в цитоплазме нейрона (трансмиссионная электронная микроскопия)
В клетках нередко обнаруживаются и другие пигменты, такие как гемоглобин, билирубин. Продукты деградации гемоглобина, депозиты железа чаще обнаруживаются в патологически измененных тканях глаза (стекловидное тело, увеальный тракт).
Дата добавления: 2015-10-21; просмотров: 114 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| КЛЕТКА И ТКАНИ | | | Интерфазное ядро |