Читайте также:
|
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
Кафедра генетики
Характеристика процесса альтернативного сплайсинга мРНК эукариот
Курсовая работа
Бакунович Александры Андреевны
студентки 2 курса,
специальность
«научно-производственная деятельность»
Научный руководитель:
кандидат биологических наук
доцент В.В. Гринев
Минск, 2015
Оглавление
Оглавление 2
Введение 3
Глава 1 Процессинг молекул првичных РНК в ядре клеток человека 4
Глава 2 Канонический и альтернативный сплайсинг РНК в клетках человека 9
2.1 Канонический сплайсинг 9
2.2 Альтернативный сплайсинг 12
2.2.1 Механизмы альтернативного сплайсинга 12
2.2.2 Процесс альтернативного сплайсинга 13
2.3 Высокая пластичность сплайсинга РНК 14
2.4 Связь канонического сплайсинга и само-сплайсинга 14
Глава 3 Факторы сплайсинга: разнообразие, строение и функции 16
Глава 4 Цис-элементы сплайсинга: разнообразие и функции 20
Глава 5 Биологическое значение сплайсинга 22
5.1 Первичные дефекты сплайсинга 22
5.2 Вторичные дефекты сплайсинга 23
Заключение 24
Список литературы 24
Введение
Процесс сплайсинга мРНК эукариот долгое время привлекает к себе внимание ученых по всему миру. Он представляет собой часть сложного многоступенчатого преобразования незрелых молекул РНК для их дальнейшей работы. В такой молекуле находятся кодирующие (экзоны) и не кодирующие (интроны) белок нуклеотидные последовательности. Функционально интроны не несут информации, необходимой для дальнейшей трансляции. В процессе эволюции у эукариот выработались механизмы для избавления от неинформативных участков. Целью их было сокращение длины и максимальное увеличение функциональности РНК. Сплайсингом называется вырезание некодирующих последовательностей из предшественника матричной РНК – пре-мРНК, – за которым следует сшивание экзонов. Этот процесс имеет большое значение для жизнедеятельности клеток и влияет на возможности экспрессии генов. Также с его помощью может увеличиваться количество белков, производимых одним геном. Это оказывает огромное влияние на процесс эволюции генов и организмов в целом.
Целью работы является характеристика процессов канонического и альтернативного сплайсинга, факторов и цис-элементов, обеспечивающих его прохождение. Также она включает в себя описание биологических эффектов, к которым могут привести нарушения в определенных стадиях его прохождения.
Глава 1
Процессинг молекул первичных
РНК в ядре клеток человека
Процессинг – совокупность преобразований предшественников РНК, которые превращают их в зрелую форму. После синтеза первичные транскрипты недостаточно активны и претерпевают ряд изменений. У эукариот процессингу подвергаются все виды РНК, у прокариот – рРНК и тРНК.
Молекулы РНК выполняют различные задачи в клетках. Они функционируют как мессенджеры для передачи генетической информации от ДНК к белкам, как основной генетический материал в клетках многих вирусов, в качестве катализаторов (например, рибозимов), имеют большое значение для синтеза белка, существуют как регуляторы экспрессии генов в живых организмах. Понимание молекулярных механизмов, посредством которых белки распознают и связывают РНК, является существенным для понимания функциональных последствий этих взаимодействий.
В зависимости от типа РНК (рибосомальная, транспортная или матричная) процессинг может проходить по-разному.
На примере матричной РНК процессинг включает следующие стадии:
1. Кэпирование
Этот процесс состоит в присоединении особой структуры, называемой кэп, на 5’-конец пре-мРНК.
С помощью экзонуклеаз от иРНК могут отделяться нуклеотиды и разрушать молекулу. Для защиты заключенной в ней информации формируется 5’-кэп. Он состоит из одного или нескольких модифицированных нуклеотидов. Его наличие является отличительной чертой эукариот. Прокариоты имеют лишь трифосфат на 5’-конце. Эта структура состоит из остатков 7-метилгуанозинтрифосфата и определенных белков. Кэпирование осуществляется с помощью РНК-трифосфатазы, гуанилтрансферазы и гуанил-N7-метилтрансферазы. Эти ферменты имеют связь с транскрипцией, участвуют в сплайсинге и процессинге 3’-конца и стимуляции трансляции. (Schoenberg D. R., Maquat L. E., 2009)
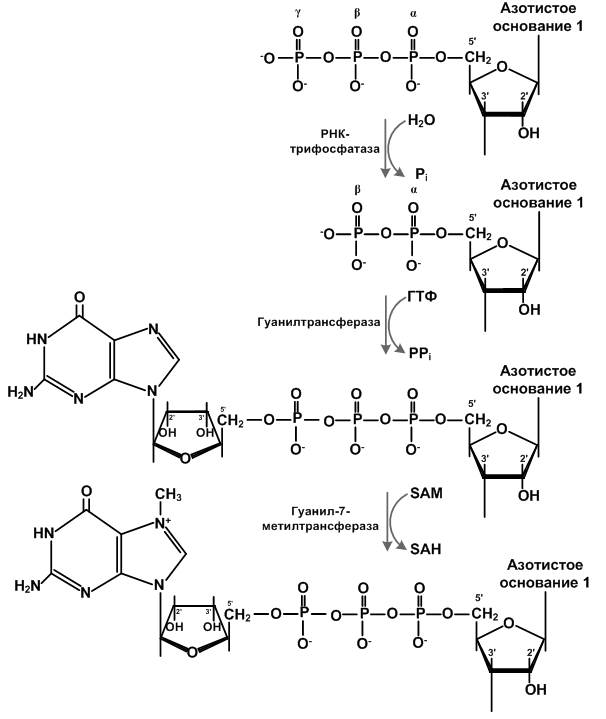
Рисунок 1.1 Механизм кэпирования
(рисунок взят с сайта https://ru.wikipedia.org/wiki/%D0%9A%D1%8D%D0%BF#/ media/File:Mechanism_of_capping.png)
2. Полиаденилирование
Процесс защиты 3’-конца пре-мРНК состоит в прикреплении большого количества молекул аденозинмонофосфата, образующих поли-А-хвост.
Азотистые основания этой структуры представлены аденином. Полиаденилирование является неотъемлемой частью процессинга эукариот. Процесс начинается после завершения транскрипции гена. Перед началом полиаденилирования мультисубъединичный белковый комплекс отщепляет 3'-концевой участок первичного транскрипта. Место расщепления определяется положением универсальных сигнальных последовательностей в первичном транскрипте. Иногда расщепление может происходить в нескольких альтернативных сайтах. Полиаденилирование даёт возможность для образования различных мРНК на основе одного гена (альтернативное полиаденилирование).
После формирования нового 3'-конца транскрипта компонент белкового комплекса – поли(А)-полимераза – осуществляет синтез поли(А)-хвоста, используя 3'-концевой нуклеотид в качестве затравки (Proudfoot N. J. et al, 2002) Поли-А-хвост защищает мРНК от разрушения, участвует в окончании транскрипции, процессах трансляции и переноса мРНК из ядра. (Hunt A. G. et al, 2008) Однако, например, у гистоновых белков эта структура отсутствует и заменена так называемой шпилькой, за которой находится гистоновый нисходящий элемент – пуриновая последовательность. (Marzluff W. F. et al, 2002)
3. Сплайсинг
Белок-кодирующие последовательности эукариотических генов (экзоны) обычно прерываются некодирующими (интронами), которые занимают большую часть длины гена. И интроны, и экзоны транскрибируются в мРНК. После этого некодирующие последовательности вырезаются из пре-мРНК через процесс сплайсинга. Только после преобразования 5’- и 3’ -концов РНК может называться зрелой. Каждый акт сплайсинга удаляет один интрон, проходя через последовательность фосфорил-транспортных реакций. Они соединяют два экзона, удаляя интрон как “лариат”. Механизм, который катализирует сплайсинг пре-мРНК, достаточно подвижен для того, чтобы охватывать все разнообразие интронов, которые присутствуют в обычной эукариотической клетке.
Присутствие крупных некодирующих последовательностей в ДНК позволяет скомбинировать экзоны различных генов. Это облегчает процесс создания новых белков. Подтверждением является факт, что многие белки современных клеток состоят из одинаковых протеиновых доменов.
Транскрипты многих эукариотических генов проходят сплайсинг в нескольких точках, позволяя одному гену производить набор различных белков. В результате РНК-сплайсинг позволяет эукариотам увеличить кодирующий по 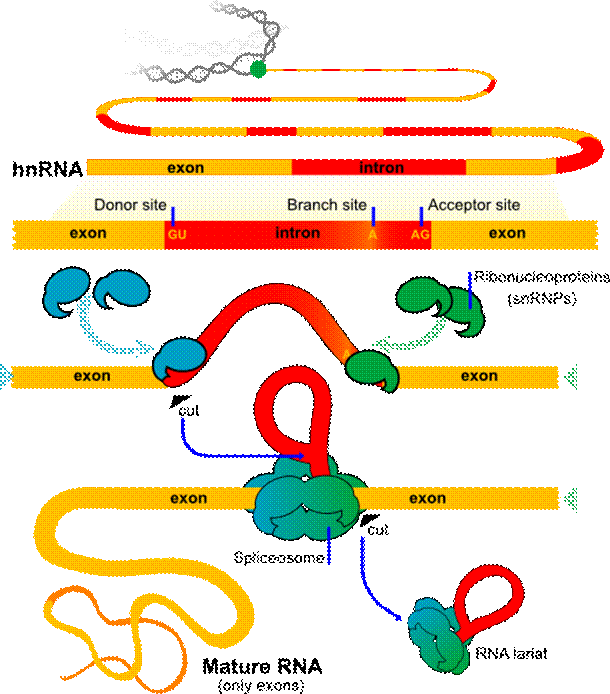 тенциал своих геномов. (Alberts В. еt al, 2007)
тенциал своих геномов. (Alberts В. еt al, 2007)
Рисунок 1.2 Схема сплайсинга
(рисунок взят с сайта http://www.clker.com/clipart-49537.html)
4. Редактирование
На протяжении процессинга преобразовываемая РНК подвергается редактированию. Оно чаще всего проходит путем химической модификации оснований и описано во всех типах РНК эукариот. Этот процесс обычно происходит в ядре клетки, цитозоле, митохондриях и пластидах.
Механизмы редактирования РНК включают в себя модификацию нуклеозидов (например, дезаминирование цитидина в уридин или аденозина в инозин) и вставки нуклеотидов без матрицы. Редактирование РНК в случае мРНК значительно изменяет последовательность кодируемого полипептида. Транслируемый белок может сильно отличаться от закодированного в ДНК генома. (Brennicke A. et al, 1999)
Редактирование РНК начинается со спаривания первичного неотредактированного транскрипта с guide RNA (ведущей РНК), которая содержит комплементарные последовательности около сайтов встраивания или удаления. Образующийся двуцепочечный участок покрывается эдитосомой – крупным многобелковым комплексом, катализирующим редактирование РНК. (Alfonzo J.D. et al, 1997) Эдитиосомный комплекс начинает встраивание уридинов по первому положению неспаренных нуклеотидов. Образуются комплементарные связи с guide RNA. Встраивание продолжается до того момента, когда в ведущей РНК встречаются A или G и останавливается при появлении C или U. (Blum B. et al, 1990)
В ходе редактирования происходит разрезание по сайту, где не образуются комплементарные пары между guide RNA и нередактированным транскриптом. Следующая стадия катализируется ферментом (концевой U-трансферазой), который добавляет U из 3’-UTP. (Simpson L., Thiemann O.H., 1995) «Открытые» концы удерживаются другими белками эдитосомного комплекса. U-специфичная экзорибонуклеаза, удаляет неспаренные уридины. Эдитосомный комплекс способен редактировать мРНК лишь в направлении от 3'-конца к 5'-концу. (Hajduk S.L., Sabatini R.S., 1998)
5. Полиметилирование
После окончания транскрипции метилтрансферазы производят ферментативный перенос метильных групп на молекулы РНК. Метилирование некодирующих РНК на 3'-конце стабилизирует эти молекулы. Наиболее распространённой модификацией является метилирование остатков аденозина по положению N6 с образованием N6-метиладенозина. Предполагается, что метилирование имеет регуляторную функцию. (Wang X. et al, 2014)
Дата добавления: 2015-09-07; просмотров: 155 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Сервитуты в ДР | | | Глава 2 |