Читайте также:
|
Жиры – основной источник энергии для синтеза АТФ. Они обеспечивают образование примерно половины энергии, потребляемой сердцем и покоящимися скелетными мышцами.
Распад жиров отличает универсальный механизм окисления жирных кислот путем последовательного отщепления от углеродной цепи двух углеродных атомов в виде ацетил-СоА. При этом образуются НАДН и ФАДH2.
Процесс окисления жирных кислот складывается из следующих этапов: активация жирных кислот, транспорт жирных кислот внутрь митохондрий, внутримитохондриальное окисление жирных кислот.
Активация жирных кислот протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А (НS-KoА) и ионов Mg2+:
R-COOH + HS-KoA + ATF ® R-CO-S-KoA+ AMF + PPi
Реакцию активизации жирных кислот катализируют три родственных фермента – ацитил-КоА-синтетазы жирных кислот, специализирующиеся на кислотах с короткими, средними и длинными цепям.
Превращение жирных кислот в ацетил-КoA происходит в митохондриальном матриксе (рис. 40). Перенос жирнокислотного остатка в митохондрии осуществляет молекула карнитина.
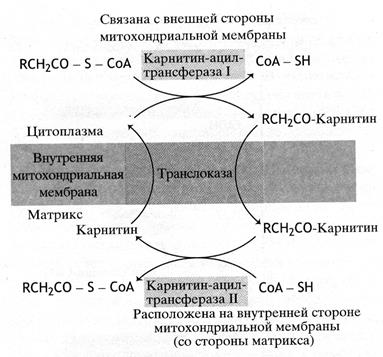
Рис. 40. Механизм транспорта жирных кислот в митохондрии
Ацильные производные карнитина проникают в матрикс, где протекает обратная реакция: ацильный остаток переносится с карнитина на тиольную группу свободного СоА, а карнитин возвращается назад за очередным остатком жирной кислоты.
Две реакции переноса ацильных групп – на карнитин и на митохондриальный СоА - катализирует два фермента: первую, протекающую на внешней стороне митохондрии, - карнитин-ацилтрансфераза I, а вторую имеющую место в матриксе, - карнитин-ацилтрансфераза II.
Внутримитохондриальное окисление жирных кислот включает четыре последовательно протекающие реакции (дегидрирование, гидратация, дегидрирование, отщепление ацетил-КоА). Реакции окисления в радикале жирных кислот происходят по b-углеродному атому, поэтому окисление называют b-окислением (рис. 41).
|
|
|
|

Рис. 41. b-окисление жирных кислот
Эти реакции называют циклом b-окисления, так как имеется в виду, что одни и те же реакции повторяются с радикалом жирной кислоты до тех пор, пока вся кислота не превратится в ацильные остатки.
Первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-дегидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3. Енол-КоА при участии фермента енол-КоА-гидратазы присоединяет молекулу воды. В результате образуется b-оксиацил-КоА (или 3-гидроксиацил-КоА), который затем дегидрируется и превращается в b-кетоацил-КоА (3-оксиацил-КоА).
Ключевой реакцией в метаболизме жиров служит реакция (тиолазная) взаимодействия КоА-SH с b-кетоацил-КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Реакция катализируется ацетил-КоА-ацилтрансферазой (b-кетотиолазой).
Образовавшийся ацетил-КоА подвергается окислению в цикле трикарбоновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь b-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА. Например, при окислении пальмитиновой кислоты (С16) повторяется 7 циклов b-окисления.
При каждом цикле образуются одна молекула NADH и одна молекула FADH2, которые отдают свои электроны митохондриальной электроннотранспортной цепи.
Окисление ненасыщенных жирных кислот идет через дополнительные стадии, включающие перемещение двойной связи в углеводородной цепи жирной кислоты и перевод этой связи в транс-форму с помощью фермента изомеразы.

Примером может служить пальмитоолеиновая кислота с двойной связью между девятым и десятым атомами углерода. Вплоть до двойной связи цепь такой кислоты укорачивается обычным окислением с образованием цис-формы. Затем изомераза сдвигает двойную связь в нужное положение и образует транс-изомер, который становится субстратом енол-КоА-гидратазы.
Дата добавления: 2015-10-23; просмотров: 132 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Переваривание и всасывание липидов | | | Биосинтез насыщенных жирных кислот |