Читайте также:
|
Содержание кислорода во внешней среде (c): 210 мл/л в воздухе и 7 мл/л в воде (при равновесии с атмосферой). Учитывая 1000-кратное различие в плотности воздуха и воды, при дыхании в воде через органы дыхания нужно перекачать массу, которая в 30000 раз больше, чем для получения того же количества в воздухе.
При дыхании в воде организовать поступление кислорода наиболее сложно, учитывая, насколько б о льшую массу нужно прокачать в сравнении с воздухом
= кислород из воздуха можно получать практически бесплатно, отсюда ясно, например, почему столь малая доля извлекается – в покое 1/7 (в легкие поступает воздух, содержащий 21% O2, уходит из них – 18%))
Жабры – принцип противотока (способ извлечь почти 100% кислорода, реально извлечь 90%)
(М) принцип противотока в биологических (например, для поддержания температуры ФЖ, с. 370–4, 398–401) и технических системах
(П) сохранение тепла за счет относительного охлаждения в конечностях и/или на поверхности тела и различие температур на поверхности тела (ФЖ, с. 336)
Количество кислорода, поступающего в органы дыхания, дает произведение Q c, где c – содержание кислорода в единице объема, Q – объем, перекачиваемый в единицу времени. Его можно рассчитать, зная геометрические характеристики системы дыхания и разность давлений между ее входом и выходом (например, по формуле Пуазейля или ее модификации для системы плоских пластин, см. формулы в справочных данных)
как пример, расчет жабр…
При наблюдаемой интенсивности обмена поток кислорода на всех этапах, предшествующих поступлению в кровеносную систему, должен быть достаточным для достижения наблюдаемой величины
Поток Q воды через жабры (представляемые как параллельные пластинки высотой h, длиной l и расстоянием между пластинами a, a << h) определяется формулой (см. справочные данные, выводится она аналогично упоминаемой выше формуле Пуазейля):
 ,
,
где D p разность давлений, которая гонит воду, h – коэффициент вязкости
Требуемый поток, позволяющий обеспечить наблюдаемую интенсивность обмена I
I = n Q c
где n – число промежутков между пластинами (которое на единицу меньше числа параллельно расположенных пластин); c – концентрация кислорода в поступающей воде.
Необходимый поток на этапе диффузии
I = D hl Dc/Dx» k D hl c/a
где D = 2,6 10–5 см2/с – коэффициент диффузии для кислорода, k – коэффициент, близкий к 1 (учитывая, что, с одной стороны, расстояние от центра потока до пластины – это a /2 и к пластине идет два потока, а с другой стороны, D c в несколько раз меньше, чем c). Для определенности далее будем считать, что k =1.
Необходимый поток на этапе поступления через поверхность обмена
I = g hl.
Ограничения на отдельных этапах имеют смысл неравенств, если их рассматривать как условия, определяющие предельно допустимые значения потока на этих этапах (и соответствующие предельные значения параметров этих этапов). Но в силу ожидаемой целесообразности организации живых организмов (нет смысла резервировать неиспользуемые возможности) естественнее их рассматривать как приближенные равенства, т.е. уравнения.
Система уравнений неполная: 3 уравнения, которые описывают связь между 4 величинами (учитывая, что во всех уравнениях фигурирует только произведение nh).
Можно дополнить эту систему еще одним уравнением, исходя из целесообразности (подумайте как именно…), а можно использовать, как неполное описание.
Из неполной системы уравнений следуют два важных количественных соотношения. Расстояние между пластинами
a = Dc/g = 0,2 мм
После этого можно преобразовать первое уравнение с учетом двух других
 .
.
Тогда можно выразить через другие параметры разность давлений, которая гонит воду через жабры
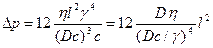 .
.
Расчет объема органов дыхания
V = a S = Dc /g I /g
(или удобно оставить как V = a I /g, учитывая, что есть численное значение a)
Если взять I из справочных данных как I 0 = 0,04 m3/4, то при m = 1 кг V = 0,04 0,2/0,33 = 24 мл. Если увеличить I до основного обмена теплокровных (т.е. увеличить почти в 20 раз), то жабры уже имеют половину объема рыбы, что, по сути практически невозможно. Тем более, невозможно увеличить до нормальной ширины метаболического диапазона. Отсюда ясно, почему рыбы не могут иметь энергопотребление как у теплокровных (еще одно ограничение в связи с тепловым балансом будет рассмотрено в следующей теме)
Развитие: аналогичный расчет легких и определение условий, при которых можно дышать ими (например, в воде, что тоже реально возможно – ФЖ, с. 267)
Для птиц на высоте – отличие от человека и других животных: однонаправленный поток воздуха (в системе дыхания с использованием легочных мешков, ФЖ, с. 71–80) – аналогично потоку в жабрах.
Тогда нет смешивания свежего и обедненного воздуха (как уменьшения в полтора раза, т.е. абсцисса кривой насыщения в принципе может быть «растянута» в те же полтора раза).
Но нужна сопутствующая физиологическая организация кровообращения. Так, даже для относительно короткой жаберной пластинки капилляр той же длины (равной длине пластинки, чтобы простейшим способом обеспечить принцип противотока) нужно рассматривать как приемлемый с точки зрения получаемого гидродинамического сопротивления.
= сюжет для чтения биологической литературы и для задания
Для сравнения: данные для геометрии сосудов кровеносной системы – ФЖ, с. 161
Число и геомерические параметры кровеносных сосудов в брыжейке собаки
| Диаметр d, мм | Число, N | Примерная длина, см | N d3 | |
| Аорта | ||||
| Крупные артерии | ||||
| Артериальные ветви | 2 400 | |||
| Артериолы | 0,02 | 40 000 000 | 0,2 | |
| Капилляры | 0,008 | 1 200 000 000 | 0,1 | |
| Венулы | 0,03 | 80 000 000 | 0,2 | |
| Крупные вены | 2 400 | |||
| Вены | ||||
| Полая вена | 12,5 |
Дата добавления: 2015-08-20; просмотров: 57 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Семинар 7. | | | Семинар 8. |