|
Читайте также: |
Энергопотребление как определяемое дыханием
(через поступление кислорода в организм животного)
Вспоминаем:
Для транспорта кислорода нужен переносчик, иначе доля полезной массы в перекачиваемой жидкости (воде) – 5 г на тонну.
Полезно проверить это значение (и другие далее – в качестве подготовки к контрольной работе):
(Д) растворимость 7 мл O2 (г)/литр при равновесии с атмосферным воздухом (21% = 150 Торр), тогда примерно 5 мл O2 (г)/литр при смешивании с воздухом легких (100 Торр после смешивания) и т.д.
Доля полезной массы значительно возрастает за счет связывания (с миоглобином или гемоглобином), но она остается относительно низкой (200–250 г на тонну – по оценке предшествующего семинара)
Это объясняет близкие зависимости энергопотребления (организованного как обеспечивающее потребление кислорода) у различных животных – основной обмен (в зависимости от массы – см. выше или в справочных данных) и метаболический диапазон (см. ниже). Но не сам вид этих зависимостей! (вернемся к обсуждению в курсе «Биофизика»)
(Д) по факту переносчик – гемоглобин (М.м 68 000 Да), который представляет собой объединение 4-х молекул миоглобина (М.м. 17 000 Да), по сути, модифицированный тетрамер миоглобина. Это означает, что он не увеличивает связывание на единицу массы перекачиваемой жидкости (в сравнении с миоглобином), а обеспечивает какой-то другой эффект.
Какой же?
Количество перенесенного кровеносной системой и использованного кислорода дает баланс поступления кислорода в кровь и его отдачи в тканях, что можно представить/выразить как разность количеств кислорода на входе и на выходе из них
 ,
,
где W – объем, перекачиваемый кровеносной системой в единицу времени, c – концентрация гемоглобина в единице объема (с пересчетом в количество переносимого кислорода при насыщении гемоглобина). Y (P) – зависимость насыщения гемоглобина от парциального давления кислорода в равновесии с кровью при контакте с внешней средой в органах дыхания (на выходе артериальной крови из них, индекс «a») и в контакте с потребляющими тканями (на выходе из них, т.е. в венозной крови, индекс «v»).
При этом от величин Pa и Pv зависят скорости поступления кислорода на других этапах (см. далее)
В силу такой зависимости эффекта переноса (кислорода связанного с гемоглобином) от DY важно не только то, сколько кислорода содержит артериальная кровь, насыщаемая при контакте с кислородом из внешней среды в органах дыхания (ее определяет насыщение Y(Pa)), но и способность отдавать кислород при давлении в капиллярах. Если величина Y(Pv) велика, то кровь будет плохо отдавать кислород.
Иными словами, для гемоглобина (как переносчика) желательно возможно б о льшее значение DY = Y(Pa) – Y(Pv) при обычных условиях.
Гемоглобин как переносчик решает именно эту задачу.
Проверяем:
(Д) кривые насыщения миоглобина и гемоглобина (БХФ, с. 104): миоглобин хорошо связывает, но плохо отдает (если в венозной крови давление не падает почти до нуля, а не может, т.к. нужно еще обеспечить диффузию к местам потребления – в митохондрии)
Миоглобин (мономер связывающего белка/переносчика) и гиперболическое связывание – плохо с точки зрения отдачи в тканях.
Для транспорта важен конечный эффект переноса на одну молекулу переносчика, который определяет разность принятого и отданного кислорода.
Для переносчика желательна сигмоидная кривая – аналогично той, которую обсуждали применительно к регуляции во Введении–1.
Вспоминаем: рассматривалось связывание фермента или активного макромолекулярного комплекса с несколькими одинаковыми молекулами (кооперативное связывание) как способ регуляции молекулярной активности.
Получаемые свойства регулируемых процессов представляют кривые связывания – зависимости скорости процесса от концентрации молекул регулятора.
Например, при кооперативном связывании регулятора (c) с ферментом (E) по схеме


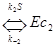
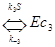 …
… 
при наличии 4-х центров связывания было получено выражение для относительной доли активной формы фермента, связанного с 4-мя молекулами регулятора (E c 4)
Y = [E c 4]/ [E] + [E c ] + [E c 2] + [E c 3] + [E c 4] =
= K4K3K2K1c4/(1 + K1c + K2K1c2+ K3K2K1c3+ K4K3K2K1c4),
которая при малых концентрациях регулятора пропорциональна 4-й степени этого регулятора и выходит на насыщение при высоких концентрациях регулятора. В этом случае вторая производная Y’’ по концентрации регулятора изменяет знак с положительного на отрицательный. Говорят, что такая кривая имеет S-образный или сигмоидный вид (подобно наклоненной и вытянутой вправо латинской букве S).
(Д) у кривой насыщения гемоглобина действительно сигмоидная форма
С чем связана сигмоидная форма?
Механизм связывания при транспорте кислорода кровью
Транспорт кислорода кровью осуществляется за счет его обратимого связывания со специальным белком – гемоглобином. В капиллярах легких гемоглобин присоединяет кислород, в капиллярах потребляющих тканей отдает его.
связывание позволяет значительно увеличить количество кислорода в объеме крови (см. далее)
При организации транспорта кислорода кровью так же, как и во многих схемах регуляции молекулярной активности, использовано кооперативное связывание. Гемоглобин имеет 4 места связывания для кислорода, т.е. максимально связывает 4 молекулы кислорода, что представляет схема.



 ,
,
где E – свободный гемоглобин, а S – кислород.
Иными словами, при регуляции (вспоминаемой выше по Введению–1) и при транспорте кровью схема связывания одинакова (при заданном числе присоединяемых молекул, которых 4 для гемоглобина) и желательные свойства кривых связывания очень похожи.
Аналогия переносчика с «идеальным переключателем»: желательно, чтобы Y(Pa) = 1 и Y(Pv) = 0.
Дата добавления: 2015-08-20; просмотров: 50 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Семинар 5. | | | Вид кривой насыщения гемоглобина |