Следовательно, в благоприятных условиях может протекать перенос энергии в направлении от цитозина к тимину.
Из сказанного также вытекает, что в фотодимеризации должны участвовать тимины, находящиеся в состоянии триплетного возбуждения. Это обстоятельство было уточнено в экспериментах с применением ацетофенона, фотосенсибилизатора, поглощение квантов излучения которым приводит к появлению значительного количества триплетно-возбужденных молекул этого сенсибилизатора. Оказалось, что облучение смеси ДНК с ацетофеноном излучением с l около 350 нм, где собственное поглощение у нуклеиновой кислоты совсем отсутствует, сопровождается накоплением тиминовых димеров в нуклеиновой кислоте. При этом следует заметить, что энергия триплетно-возбужденных состояний ацетофенона достаточна для ее передачи на тимин, но не позволяет возбуждать другие азотистые основания.
Тушители триплетно-возбужденных состояний (например, О2) способны существенно ослабить процесс фотодимерации тимина в растворе этого соединения. Однако на фотодимеризацию смежных остатков этого основания в составе ДНК кислород не влияет. Это связано с тем, что смежным тиминам в составе ДНК не требуется сближение путем диффузии для образования димера, тогда как для тушения кислород должен приблизится к триплетно-возбужденному тимину диффузионным путем. Интересно, что ряд встраивающихся в двойную спираль ДНК акридиновых красителей эффективно защищают нуклеиновую кислоту от повреждения УФ излучением. Наиболее эффективен в этом плане краситель акрихин. Данное действие этих красителей связано с их способностью тушить триплетные возбуждения оснований нуклеиновой кислоты за счет переноса энергии возбуждения на молекулу красителя.
Хотя фотодимеризация тимина – основная причина повреждения ДНК при действии УФ излучения, в этой нуклеиновой кислоте возможны и другие фотоиндуцированные реакции. Например, известно, что под влиянием излучения могут формироваться димеры типов Ц-Ц или Т-Ц.
Еще один тип фотохимических реакций в нуклеиновых кислотах - фотогидратация пиримидиновых оснований. В результате из цитозина и урацила образуются соответствующие 6-окси-5-гидроксипроизводные:
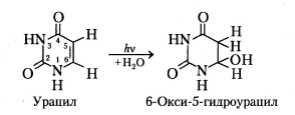
Показано накопление таких производных при действии УФ излучения на растворы цитозина и урацила, на РНК и однонитевую ДНК. В двухнитевой ДНК данные производные, однако, не выявляются. Тушители триплетных возбуждений на скорость накопления 6-окси-5-гидроксипроизводных цитозина и урацила влияния не оказывают, что позволяет предположить, что реакция фотогидратации протекает через состояние синглетного возбуждения указанных оснований. В отличие от фотодимеризации реакция фотогидратации необратима – разрушения ее продуктов при действии излучения не происходит. Однако 6-окси-5-гидроксипроизводные урацила и цитозина термически неустойчивы и распадаются при нагревании с образованием исходных соединений.
УФ излучение может вызвать также формирование ковалентных сшивок между молекулами ДНК и белками. В условиях эксперимента показано также, что облучение совместных растворов цитозина или урацила и ряда аминокислот (серина, цистина, метионина, лизина, аргинина, гистидина, триптофана, тирозина и фенилаланина) приводит к формированию ковалентных связей между аминокислотами и 5-м или 6-м углеродными атомами пиримидиновых оснований. В частности, формировался 5-S-цистеин-гидроурацил. Иногда УФ излучение вызывает формирование ковалентных сшивок между отдельными молекулами ДНК.
Если дозы облучения велики, в нуклеиновых кислотах накапливаются разрывы полинуклеотидных цепей. В двухцепочечных молекулах гораздо чаще (в 100-1000 раз) возникают однонитевые разрывы. Однако вклад даже таких разрывов в биологические эффекты излучения, связанные с нуклеиновыми кислотами невелик.
Повреждение генома клеток УФ излучением запускает еще один очень значимый фотобиологический процесс – фотореактивацию. Этот эффект заключается в частичном восстановлении поврежденного УФ излучением генома после дополнительного облучения клеток в более длинноволновом спектральном диапазоне. Для бактерий эффект фотореактивации УФ-индуцированных повреждений генома характеризуется спектром действия в диапазоне от 300 до 500 нм с максимумом при 380 нм.
Фотореактивация определяется наличием в клетках специального фермента, стимулирующегося фотореактивирущим излучением. Интересно, что сам по себе этот фермент данное излучение не поглощает. Хромофорная группа образуется при взаимодействии этого протеина с циклобутановыми димерами пиримидиновых оснований. Поглощение квантов видимого света таким комплексом приводит к распаду циклобутановых димеров и регенерации исходных оснований.
Эффект фотореактивации наблюдается и в эукариотических клетках. Показано, например, что фотореактивация генома лимфоцитов и фибробластов человека наблюдается при облучении в области 300-600 нм. Максимум спектра действия в этом случае находится при 400 нм.
Дата добавления: 2015-07-25; просмотров: 92 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Действие ультрафиолетового излучения на нуклеиновые кислоты. | | | Учебное пособие |