Читайте также:
|
Выше упоминалось о связях трех типов (вещественных, энергетических и информационных) между компонентами экосистемы. Их изученность на разных уровнях подразделения по элементам еще очень недостаточна. Исторически прежде всего начали изучать вещественно-энергетические взаимосвязи в рамках классической трофодинамики и моделей типа «хищник-жертва». Кроме того, развивались представления о совокупностях отдельных групп организмов, связанных общностью местообитания. Последнее направление особенно интенсивно развивается в рамках геоботаники - науки о растительном покрове Земли, распространении и закономерностях размещения растительных сообществ, а особенно в фитоценологии - разделе геоботаники, изучающем растительные сообщества: их классификацию, состав, смену, изменения; взаимоотношения между собой и между компонентами сообществ в них и в окружающей среде.
Некоторые черты сходства с геоботаническими концепциями можно усмотреть также в гидробиологических работах, посвященных бентосным сообществам. И в том и в другом случае исследователи имеют дело с комплексами преимущественно одного трофического уровня. Поэтому говорить о непосредственных энергетических связях между ними не приходится. Что касается косвенных (конкурентных) взаимодействий, то в ряде случаев было показано, что такая система стремится к их минимизации. Такие же выводы, касающиеся наземной растительности, появились позже.
В последнее время выясняется очень большая роль трофических связей так называемого нехищного типа, которые особенно характерны для низших организмов водной среды. Эти связи осуществляются путем выделения в раствор различных метаболитов. Несомненно, что такого рода «экологический метаболизм» имеет место в почвах наземных экосистем.
Таким образом, в экосистеме можно выявить связи разной природы. Исследователи стремятся к анализу все новых и новых типов связей (например, сенсорных). Это, с одной стороны, открывает новые интересные перспективы, а с другой - все более проблематичной становится возможность отыскания неких универсальных единиц для измерения силы взаимодействия организмов друг с другом в рамках целой экосистемы.
С одной стороны, к такого рода оценкам можно подойти через информационные показатели зависимости между компонентами, с другой - через определение энергетических последствий тех или иных взаимодействий («энергия как валюта»). Видимо, возможно совмещение этих двух подходов. Однако во всех случаях на окончательный результат большое влияние будут оказывать характер наших предварительных допущений, степень генерализации, принципы выделения элементов системы и другие факторы, в сильной мере зависящие от биологической интуиции и опыта исследователя.
Термины «эффективность функционирования», «стабильность» и «степень организованности» не имеют однозначного определения, когда они касаются экосистем или их крупных блоков, однако в последнее время эти понятия все чаще обсуждаются.
Для выяснения эффективности функционирования, видимо, необходимо допустить наличие «щели» у системы (телеономический подход). Поскольку такие допущения могут быть различны, то и оценки эффективности функционирования также будут разными.
Говоря о живых организмах, часто предполагают их стремление к максимизации биомассы, которое в крайних случаях приводит к так называемым экологическим взрывам. В то же время подчеркивается их «стремление» к сохранению устойчивости, структурной организованности в условиях хаосогенной среды. Однако и та и другая «цели» требуют соответствующих энергетических затрат, и, таким образом, в этом плане они конкурируют.
Большинство экологов считают разнообразие трофической сети, состоящей из многих трофических цепей, выраженное в информационных единицах (на базе оценки вероятности переноса энергии по тому или иному каналу), мерой стабильности сообщества (модели Р. Макартура и т. п.). Близка к этому и такая же оценка статической картины «видового» или «группового» разнообразия (через вероятность распределения биомассы или энергии по видам или группам сообщества). Оценка разнообразия будет, как и во многих других случаях, зависеть от числа и характера выделенных элементов экосистемы.
Имеющиеся формальные критерии организованности можно, в принципе, использовать при описании и сравнении различных моделей экосистем. Однако таких критериев недостаточно. С биологической точки зрения не всегда упорядоченность (структурная негэнтропия) прямо тождественна организованности, если иметь в виду (а биолог всегда имеет это в виду) эффективность выполнения жизненно важных функций. При формальной оценке упускаются многие критерии, определяемые пока только качественно.
Известно, что при прочих равных условиях (отношение поверхности к массе, температура среды и т. д.) метаболизм покоя выше у более высокоорганизованных живых существ. Это показано при сопоставлении крупных филогенетических групп организмов (типов, классов) и при более дробном делении. Так, например, установлено, что обмен веществ у более высокоорганизованных плацентарных млекопитающих выше, чем у сумчатых. Это явление объясняется тем, что поддержание высокоорганизованной структуры требует дополнительных «вложений» энергии для противодействия закону организационной деградации.
Касаясь оценки степени организации на уровне сообщества, следует говорить о суммарном (аддитивном) потоке тепла на единицу поверхности за единицу времени. Тогда поток тепла (непревратимой энергии) при прочих равных условиях и будет характеризовать степень организованности. Другими словами, чем более организован биоценоз (и соответствующая экосистема), тем большую энергетическую плату он должен отдавать на поддержание этой организованности.
Отсюда следует, что высокоорганизованное сообщество вынуждено минимизировать энергетический вклад в накопление биомассы. Это в некоторой степени противоречит тенденции максимизировать биомассу, проявляющуюся в потенциально возможных вспышках размножения и экспансиях в свободные ниши.
Выше разбирались некоторые подходы к разделению экосистемы на блоки, выделению связей и к оценке этих особенностей. Такой анализ учитывал прежде всего участие разных компонентов в трансформации и передаче вещества, энергии и информации. Однако по отношению к наиболее сложным биотическим компонентам таких подходов недостаточно. Для познания биоценозов обязательно требуется рассмотрение их в разных аспектах. При этом множество организмов экосистемы (общее число особей, биомасса, продуктивность и т. д.) как бы разбивается на различные группы (подмножества) по тому или иному признаку. Такие процедуры в самом общем смысле нередко именуют «анализом структуры».
При конкретных исследованиях наземных экосистем или их крупных блоков (растительность, животное население и т. п.) всегда осуществляется таксономический анализ, т. е. указывается название и число видов, семейств и других систематических категорий. Простейший вариант - составление списка имеющихся форм организмов и их таксономических групп. На следующих этапах на базе количественных учетов в разное время и знания различных экологических и физиологических параметров (индивидуальная биомасса, темпы роста, размножения, смертность, активность, энергетика и химизм метаболизма и т. д.) анализ дополняется соответствующими количественными показателями (численность, биомасса, продуктивность, участие в потоке энергии и круговороте вещества и т. д.). В настоящее время ученые только исследуют (применительно к наземным экосистемам) этот второй этап, опираясь на методический опыт и факты целого ряда биологических и географических наук. До сих пор еще нет полных сведений о видовом составе, обилии и вещественно-энергетических трансформациях в какой-либо наземной экосистеме. Можно сказать, что пока еще не существует ни одного квадратного метра суши, изученного хотя бы в отношении состава и обилия видов всех населяющих его организмов. Имеется лишь сравнительно небольшой эмпирический материал гораздо большей степени генерализации.
По такой же схеме может проводиться не только таксономический, но и другие анализы, группирующие биотические компоненты по иным признакам. В частности, большой интерес с точки зрения филогенеза на уровне сообщества представляют ареалогический (по признаку принадлежности к тому или иному типу ареала) и географо-генетический (по признаку общности территории возникновения) анализы. Представляется весьма перспективным анализ, показывающий характер распределения организмов по признаку индивидуальной биомассы (размера). Интерес такого подразделения определяется, в частности, тем, что по размерным признакам можно сравнивать любые, даже самые далекие по облику и происхождению биоценозы. Помимо этого, индивидуальные размеры связаны с энергетикой известным уравнением

где М - интенсивность метаболизма в данных условиях, А -индивидуальная масса организма, k и b - коэффициенты. Это также показывает большую общность, в принципе допустимую при проведении размерного анализа разных биоценозов.
Познание трофической структуры биоценозов - главнейший этап на пути к проникновению в функциональную сущность различных компонентов экосистем. Естественно, проблема эта разрабатывалась преимущественно экологией животных, так как зеленые растения, с которыми в основном имеют дело геоботаники и экологи растений, функционально однородны, принадлежат к одному трофическому уровню. Многие принципы гидробиологии могут быть успешно использованы и на суше, хотя специфика наземных сообществ создает ряд дополнительных проблем.
Энергетический принцип изучения трофических взаимоотношений привел к возникновению понятия о трофических уровнях. Это понятие близко к представлениям о цепях питания и пирамидах биомасс. Как уже указывалось, к первому уровню относятся автотрофы, ко второму - гетеротрофы первого порядка, к третьему - хищники первого порядка и т. д. Таким образом, в энергетическом плане сообщество разделяется на ряд соподчиненных ярусов (рис. 3). Это же отчасти отражается и в пространственном расположении организмов, соответственно преломляясь в гравитационном поле Земли в направлении потока солнечной радиации.
Выделение трофических уровней при анализе структуры экосистемы - далеко идущая идеализация в анализе структуры экосистем. Особь одного и того же вида животных может быть одновременно членом нескольких уровней. При этом приходится ориентироваться на преобладающие черты трофики или же соответствующие части энергетического бюджета популяции относить к разным уровням. Например, особи в основном растительноядных видов (некоторые грызуны, копытные и т. п.) могут поедать и животную пищу. Обычно доля ее крайне незначительна. Поэтому данные группировки животных можно целиком относить ко второму трофическому уровню. В других случаях доля животной пищи более ощутима. Так, многие птицы в течение года примерно половину своих пищевых потребностей удовлетворяют за счет животного, а другую половину - за счет растительного корма. Такие животные вносят свой «энергетический вклад» в формирование второго и третьего трофических уровней поровну. Могут встречаться и еще более сложные ситуации.
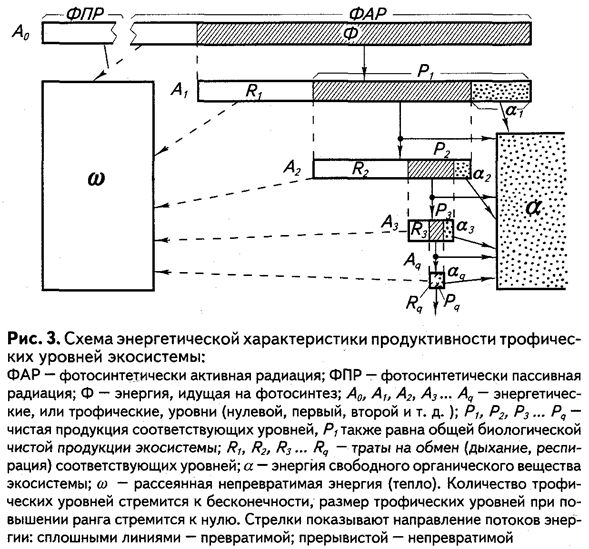
Касаясь этого вопроса, Г.Г. Винберг, однако, пишет: «Весьма просто подвергнуть критике представление о трофических уровнях и указать на многие трудности, встречающиеся уже при отнесении гидробионтов к тому или другому уровню. Не всегда легко даже провести границу между автотрофными и гетеротрофными организмами. Как известно, некоторые способные к фотосинтезу перидиниевые водоросли могут в то же время заглатывать диатомей, мало уступающих им по размерам... Легко привести и другие примеры, но они колеблют безусловную необходимость и полезность представлений о трофических уровнях не более, чем невозможность реального движения без трения колеблет основные законы механики. Другое дело, что концепция трофических уровней не передает все существующие стороны трофических взаимоотношений и требует конкретизации и дополнений»1.
Разделение биоценоза на конкретные трофические группы должно отражать сходство и различие характера воздействия организмов на вещественно-энергетические трансформации внутри экосистемы. Изучая структуру биотического сообщества в плане трофических уровней, мы абстрагируемся от конкретного строения и структуры имеющихся взаимосвязей, поскольку главное в этом случае — выяснить приход и расход основных энергетических ресурсов экосистемы в том виде, в каком этот баланс естественно сложился на каждом из уровней. Однако часто необходимо обратиться к конфигурациям конкретных взаимосвязей.
Эти конфигурации сильно зависят от степени пищевой специализации конкретных организмов биотической системы. Так, преобладание малоспециализированных трофических групп резко увеличивает число каналов взаимодействия и энергетических потоков даже при одинаковом соотношении трофических уровней. Это, в свою очередь, оказывает решающее влияние на характер регуляции основных функций экосистемы. Следовательно, для познания функционирования биотической системы необходима как схематизация явлений с применением концепции трофических уровней, так и конкретизация их до трофических групп, особенно тех полифункциональных блоков экосистемы, которые входят сразу в несколько трофических уровней. Более мелкие трофические группировки даже внутри одного уровня зачастую отличаются существенной функциональной неоднородностью (например, фитофаги и сапрофаги внутри второго трофического уровня).
Разумеется, воздействия на вещественно-энергетические трансформации могут осуществляться опосредованно, минуя чисто трофические отношения. Наиболее яркий пример этому - воздействие механической работы некоторых животных. В ряде наземных экосистем большую землеройную работу выполняют муравьи, дождевые черви, мокрицы, термиты, кроты, слепушонки, сурки, суслики, лемминги, пеструшки, луговые собачки, карманчатые гоферы, вискачи, некоторые змеи и черепахи и даже птицы (буревестники, чистики, ракшеобразные и ряд воробьиных).
Большое значение может иметь механический перенос семян, спор, яиц, цветочной пыльцы, а также паразитов, бактерий и вирусов. К воздействиям такого же рода следует отнести антибиоз, образование растениями плотных поселений, дернин, затененных участков и т. п. Такие влияния, имеющие свою специфику, удобнее подробно анализировать отдельно от трофических отношений. При этом даже косвенное действие на вещественно-энергетические (материально-энергетические) трансформации - чаще всего суть воздействия на трофические связи. Ибо питание - это пополнение энергетических и вещественных ресурсов, затраченных в процессе жизнедеятельности. Так, те же землерои, изменяя свойства почвы, влияют на условия произрастания растений и т. п.
По типу питания (включая тип источников энергии и определенных веществ) все существующие организмы делятся на следующие группы: 1) хемоавтотрофы; 2) фотоавтотрофы; 3) хемогетеротрофы; 4) фотогетеротрофы. Наземные экосистемы содержат преимущественно (а иногда и исключительно) представителей второй и третьей из указанных групп. Выделенные четыре крупные трофические группы мы предложили условно называть типами функционально-трофических групп.
Тип фотоавтотрофов в функциональном отношении представляется весьма однородным. Все организмы этого экологического типа используют в качестве источника энергии солнечное излучение, в качестве источников углерода, азота и других химических элементов - неорганические соединения (углекислый газ и минеральные соли). Фотоавтотрофы вместе с хемоавтотрофами (хемосинтезирующими организмами) образуют первый трофический уровень.
Наиболее разнообразен во всех отношениях функционально-трофический уровень хемогетеротрофов. Всех организмов этого типа объединяет то, что в качестве источника энергии они могут использовать лишь химическую энергию органических веществ. Эти же вещества необходимы и как источник углерода. Тип хемогетеротрофов включает в себя все трофические уровни, начиная со второго. Сюда относятся животные, большинство бактерий, грибы, актиномицеты, немногие бесхлорофилльные высшие растения. По числу видов этот тип превосходит во много раз все остальные, вместе взятые. Внутри типа хемогетеротрофов намечается ряд иерархически соподчиненных более мелких групп, отличающихся по характеру питания и, следовательно, по своим функциям в материально-энергетических процессах биотических систем.
При такой классификации хемогетеротрофов весьма часто в одну группу целиком входят крупные систематические категории, единые в трофическом отношении. Однако нередко даже один и тот же вид резко меняет характер питания (переключается в принципиально иные отделы трофической сети): по сезонам, на разных стадиях онтогенеза, или, реже, в отдельные годы. Более того, иногда питание особей разного пола на одной стадии развития и в одно время резко различается. Очень часто разнятся по трофике личинки и имаго насекомых. У многих двукрылых кровососы - только самки; многие птицы в зависимости от возраста и сезона переходят от животной пищи к растительной и наоборот.
Все это при классификации трофических функциональных групп заставляет дифференцированно подходить к оценке видов и систематических категорий. При этом, если главное внимание уделять не филогенетической близости, а сходству и различию трофики, зачастую даже один и тот же вид (в зависимости от сезона, пола, стадии, развития) должен быть отнесен к разным функционально-трофическим категориям. Поэтому даже при самом дробном подразделении экологическая, функционально-трофическая классификация сомкнется с филогенетической лишь отчасти.
Рассмотрим подробнее классификацию типа хемогетеротрофов. Выявление внутри этого самого сложного типа соподчиненных категорий особенно необходимо. Тип делится на два функционально-трофических класса: 1) прототрофов; 2) органитрофов.
Прототрофы включают большую часть грибов и актиномицетов, а также некоторые небольшие группы бактерий. В качестве источника энергии и углерода эти организмы используют, как это свойственно хемогетеротрофам, органические соединения. Азот же для построения белков они могут усваивать в виде неорганических солей (как и фотоавтотрофы) или в ряде случаев поглощать из атмосферы.
Органитрофы для получения энергии и всех основных веществ используют органические соединения. Этот класс объединяет всех животных, большую часть бактерий и паразитические растения. Подробное подразделение класса органитрофов особенно важно ввиду функционального разнообразия его представителей.
Необходимо сделать несколько предварительных замечаний. При разделении экосистемы на трофические уровни, о чем шла речь, выясняются общие особенности последовательной передачи и рассеивания энергии первичной продукции. В этом случае можно соответствующие части энергетического бюджета некоторых видов и групп относить к разным трофическим уровням. Такая схематизация, с одной стороны, отчетливее выявляет сущность макроэнергетических процессов экосистемы, показывает расход и приход «горючего», энергия которого двигает биотические процессы. С другой стороны, одновременно с этим затушевывается картина взаимного сплетения различных по «размеру поперечных сечений» и по скорости потока энергетических, химических и информационных каналов, каналов взаимосвязи. Поэтому при более дробной классификации по функционально-трофическому признаку организмы со смешанным характером питания следует выделять в отдельные группы. Такие полифагийные организмы существенно усложняют структуру пищевой сети, выполняют одновременно несколько функций и могут иметь важное регуляторное значение для больших отделов этой сети.
Исходя из указанных предпосылок, класс органитрофов мы делим на следующие разделы (отряды): 1) биофагов; 2) сапробиофагов; 3) сапрофагов. Первые питаются тканями и соками живых организмов, вторые и третьи - мертвым органическим субстратом. К сапрофагам относят и сапрофитов (бактерий), и сапрозоев, понимая под этим словом широкий круг различных животных и растений, питающихся отмершими остатками на разных стадиях разложения, освобождая при этом неорганические вещества. Сапробиофаги промежуточны, так как потребляют и живую и мертвую органическую материю. Разряд сапрофагов объединяет большинство бактерий и многих беспозвоночных животных. Особенно многочисленны представители разряда в богатых гумусом почвах и в подстилке. К сапробиофагам, так же как и к разряду биофагов, относят преимущественно животных.
Дальнейшее разделение (или объединение) идет по функционально-трофическим семействам. Так, сапрофаги объединяют семейства детритофагов, копрофагов, некрофагов. Биофаги включают представителей хлорофитофагов (потребителей зеленых растений), ахлорофитофагов (потребителей незеленых растений), зоофагов, зоофитофагов. Семейства далее распадаются на функционально-трофические трибы. Например, хлорофитофаги содержат следующие трибы: филлофагов, ризофагов, ксилофагов, карпофагов, антофилов;


Рис. 4. Схема энергетических связей в трофической сети биотического сообщества (с детальностью до некоторых функционально-трофических семейств).

 Заштрихованные блоки — отмершая биомасса и соответствующая ей энергия. Отражены лишь энергетические каналы. Если построить также каналы взаимовлияний, то в системе «потребитель потребляемый» в случае потребления энергии живой биомассы чаще выявляется отрицательная обратная связь, в случае потребления энергии отмершей биомассы — положительная обратная связь. Однако всегда сила и характер связи в таких «замкнутых контурах» меняются во времени даже на одних участках трофической сети (метастабильное состояние)
Заштрихованные блоки — отмершая биомасса и соответствующая ей энергия. Отражены лишь энергетические каналы. Если построить также каналы взаимовлияний, то в системе «потребитель потребляемый» в случае потребления энергии живой биомассы чаще выявляется отрицательная обратная связь, в случае потребления энергии отмершей биомассы — положительная обратная связь. Однако всегда сила и характер связи в таких «замкнутых контурах» меняются во времени даже на одних участках трофической сети (метастабильное состояние)
Изучение конфигурации пищевых сетей (см. рис. 4) в течение всего деятельного периода выявляет внутригодовую динамику «структурной формулы» биотического сообщества. Многолетние циклы таких колебаний с течением геологического времени приобретают определенный поступательный характер, что приводит к изменению свойств этой «формулы», а также к изменению биоценоза и к филогенетическим преобразованиям его взаимосвязанных компонентов. Историческое развитие захватывает не только отдельные организмы, но и всю их совокупность.
Дата добавления: 2015-10-16; просмотров: 137 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| РАЗДЕЛЕНИЕ ЭКОСИСТЕМЫ НА БЛОКИ | | | ФАКТОРЫ РАСПРОСТРАНЕНИЯ ОРГАНИЗМОВ |