Читайте также:
|
Основой этого вида памяти являются структурные изменения в нейронах, длительность (часы, дни и на протяжении всей жизни при повторении информации), ее объем практически безграничен. Долговременная память по своему механизму качественно отличается от кратковременной и промежуточной памяти, так как не нарушается при таких экстремальных воздействиях на мозг, как механическая травма, электрошок, наркоз и т.д. Долговременная память формируется с помощью кратковременной и промежуточной памяти, при этом важную роль играют синаптические процессы.
Синаптические процессы. Под влиянием обучения в ЦНС образуются новые синапсы, увеличиваются их размеры, количество, количество медиаторов, разрастаются дендриты, увеличивается число шипиков на них, количество коллатералей аксонов – все эти явления в течение месяца исчезают. Различные медиаторы могут оказывать разные эффекты в процессах усвоения и хранения информации. Серотонин, в частности, ускоряет обучение и удлиняет сохранение навыков при положительном эмоциональном подкреплении, например пищевом, и блокирует у животных выполнение оборонительных навыков. Норадреналин ускоряет обучение в условиях применения отрицательного подкрепления (электрокожного).
Моноаминергические механизмы, связанные с подкреплением, активируют внутриклеточные постсинаптические процессы с участием циклических нуклеотидов – цАМФ и цГМФ. В результате последующих метаболических внутриклеточных процессов синтезируются специальные белковые молекулы, которые, в свою очередь, стабилизируют первичные изменения синаптических мембран. В результате этого в структурах мозга формируются зоны повышенной синаптической проводимости, что и определяет формирование соответствующих энграмм памяти.
Многочисленные эксперименты показали, что связывание индифферентного возбуждения с безусловным при выработке условных рефлексов может быть результатом синаптического облегчения реакций нейрона на индифферентный стимул в ходе сочетаний индифферентного стимула с безусловным подкреплением (Л. Л. Воронин, Е. Н. Соколов, X. Матисс и др.). Соответствующий феномен получил название гетеросинаптического облегчения (фасилитации). В чем оно конкретно проявляется?
Будем проводить электрическую стимуляцию афферентных проводников, конвергирующих к одному и тому же нейрону. При слабом раздражении первого проводника («условный» стимул) нейрон отвечает небольшим ВПСП. При сильном раздражении второго проводника («безусловный» стимул, подкрепление) нейрон отвечает ВПСП большой амплитуды, на фоне которого генерируются потенциалы действия. Как выяснилось, сочетание первого и второго воздействий в течение нескольких минут постепенно приводит к усилению амплитуды ВПСП на «условный» стимул и возникновению потенциалов действия, аналогичных тем, которые возникали в ответ на «безусловный» стимул. Облегчение ответов на «условный» стимул в таких опытах сохранялось 10 – 40 мин после отмены «безусловного» стимула, т.е. характеризовалось следовым эффектом. Последний можно было трактовать как проявление запоминания информационно значимых (условных) раздражителей, поступающих к регистрируемому нейрону.
Т.Е. Греченко и Е.Н. Соколов выполнили эксперименты по выработке на изолированной соме нейрона неассоциативных и ассоциативных форм обучения, где использовались различные варианты ассоциирования стимулов. Например, в качестве условного подпорогового стимула служила микроаппликация медиатора (ацетилхолина или серотонина), в качестве безусловного стимула (эффективного относительно генерации потенциала действия) – внутриклеточный деполяризационный импульс тока. После неоднократных сочетанных воздействий применение подпороговой дозы ацетилхолина или серотонина вызывало возникновение ПД. Эффект длился у разных нейронов 5 – 40 мин и более.
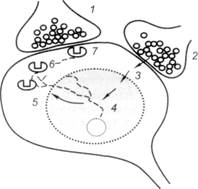
Рис. 5.9. Механизм повышения эффективности синаптической передачи в процессе образования временной связи между условным и безусловным возбуждением в ходе выработки условного рефлекса (по X. Матиссу)
Наиболее значимые результаты удалось получить в исследованиях с нейронами гиппокампа где следовые изменения синаптической активности в процессе переработки входной импульсации сохранялись на протяжении часов и даже дней.
Каковы причины, лежащие в основе тетеросинаптического облегчения? Предполагается следующий принципиальный механизм. Синаптический контакт-1 на некотором интеграторном нейроне проводит возбуждение от условного раздражителя (рис. 5.9). Исходно нейрон на условный раздражитель не реагирует из-за низкой реактивности постсинаптической мембраны синапса-1 к выделяемому медиатору-1. Однако под влиянием квантов медиатора здесь возникают кратковременные конформационные перестройки постсинаптической мембраны.
Синаптический контакт-2 обеспечивает передачу возбуждения на интеграторный нейрон от безусловного раздражителя. Выделяемый здесь медиатор-2 вызывает выраженную ответную реакцию. По-видимому, возбуждения безусловного происхождения помимо чисто электрических эффектов возбуждения нейрона-мишени ускоряют протоплазматические биохимические реакции (3), направленные на активацию генома (4) ядра нервной клетки. Результатом такой активации является изменение синтеза РНК и синтеза в рибосомах нейрона (5) нейроспецифического белка. В дальнейшем вновь синтезированный белок перемещается (6) в сторону постсинаптической мембраны того синапса, который подвергся подпороговой активации в результате воздействия на него возбуждения условного происхождения. На заключительном этапе происходит встраивание белковой молекулы в постсинаптическую мембрану данного синапса (7), при этом функциональная активность синапса-1 изменяется: из неэффективного синапс превращается в эффективный. Процесс обучения ухудшается при ингибировании синтеза РНК.
Интересная гипотеза, объясняющая процесс трансформации функциональных свойств неэффективных синапсов, была разработана И.П. Ашмариным (1975), который предположил, что в результате непрерывного поступления к нейрону сигнальной информации в протоплазме нейрона усиливается синтез специфических белков-антигенов (гликопротеинов памяти), характерных для данного нейрона. Эти белки являются компонентами синаптических мембран и (когда нейрон пребывает в состоянии относительного покоя) синтезируются в количествах, достаточных только для их обновления. В период повторной импульсации, например в ходе реверберации, при выработке ассоциативной связи происходит усиление синтеза белков-антигенов, при этом синапс из неэффективного переходит в разряд эффективных, т. е. начинает «узнавать» первичный раздражитель, обеспечивает его облегченное проведение через нейрон.
Важную роль в консолидации памяти играют нейропептиды. Известно, что пептиды могут находиться в пресинаптических терминалях в качестве сопутствующего медиатора. Например, вместе с норадреналином часто выделяется нейропептид У, опиоидные пептиды, соматостатин. Дофамин часто выделяется окончаниями аксонов вместе с холецистокинином, энкефалином; ацетилхолин – с вазоактивным интестинальным пептитодом (ВИП), энкефалином, люлиберином; серотонин – с веществом Р, тиреолиберином, холецистокинином (И.П. Ашмарин). Нейропептиды оказывают пре- и постсинаптическое модулирующее действие. Доказано, что выделение пептидов в пресинаптических окончаниях зависит от частоты работы нейрона, при этом избыточное выделение пептида-спутника всегда наблюдается при усилении активности нейронов.
Как влияют пептиды на клетку-мишень? Пептид-спутник может значительно повысить сродство рецептора постсинаптической мембраны к основному медиатору. Например, ВИП усиливает сродство к ацетилхолину более чем в 10000 раз.
Пептиды выделяются нервными клетками не только в синаптическую щель, но и во внеклеточное пространство. Эффективный путь диффузии пептидов может составить 1 мм и более. Этого вполне достаточно, чтобы охватить своим влиянием (непосредственно или через систему вторичных посредников – циклических нуклеотидов, ионов кальция) микроансамбль нейронов, изменяя его суммарную возбудимость, синхронизируя реактивность его нейронов, обеспечивая тем самым их включение в некоторый адаптивный поведенческий акт. Очевидно, мозаика образующихся функциональных групп нейронов, подвергнутых нейрохимическому воздействию пептидов, может быть одним из механизмов оперативного функционального объединения нервных клеток в ходе обучения, в явлениях памяти (Г. А. Вартанян, А. А. Пирогов, 1991, А. Мокрушин, М. Самойлов, 1999).
Прямые доказательства участия пептидов в механизмах памяти можно получить, контролируя их выделение в экстраклеточное пространство по показателям изменения химического состава цереброспинальной жидкости у животных при обучении. Разработка этого направления серьезно ставит вопрос о возможности переносов выработанных навыков от одного животного к другому (Г. Унгар) с помощью образованных в ходе обучения и «отвечающих» за соответствующий навык специфических пептидов. Гипотеза Г. Унгара основана на фактах накопления в мозге особого пептида, кодирующего страх темноты и способного транспортировать этот страх от животных – обученных доноров к необученным реципиентам. Этот пептид – скотофобин (от греч. «боязнь темноты») был синтезирован и использовался в различных лабораториях для проверки способности переноса химическим путем определенной информации. Было установлено, что скотофобин действительно вызывает стрессовую реакцию у животных, когда они находятся в темноте. При выработке у крыс рефлекса избегания темноты количество скотофобина возрастает в первые 6 дней обучения, затем постепенно снижается и через 15 дней обнаружить его в мозге не удается. Несомненный интерес для выяснения роли пептидов в процессах памяти представляют опыты с АКТГ (адренокортиктропным гормоном), меланостимулирующим гормоном, вазопрессином, позволяющие судить о их способности стимулировать запоминание при введении извне (И. П. Ашмарин, Р. И. Крутиков). Некоторые фрагменты АКТГ участвуют в механизмах сохранения памяти. Удаление у животных гипофиза существенно нарушает проявление ранее выработанных оборонительных навыков.
Установлено, что животные с врожденным дефектом выработки вазопрессина, не способны к образованию оборонительных навыков. Выработка навыка восстанавливается у этих животных только при дополнительном введении им вазопрессина. Характерно, что при этом у животных страдает не сам процесс обучения, а именно консолидация сформировавшихся энграмм.
Другой олигопептид – окситоцин – нарушает сохранение выработанных навыков независимо от типа обучения животных.
Установлено, что нейропептиды в механизмах памяти тесно взаимодействуют в синапсах с медиаторами и что после обучения кругооборот катехоламинов в мозге увеличивается.
Заметно улучшают память эндогенные опиатные пептиды – эндорфины и энкефалины.
Роль синтеза белка. После открытия способа кодирования генетической информации в ДНК (генетической памяти) и успешного изучения иммунологической памяти были предприняты попытки отыскать молекулярные основы нейронной памяти, т. е. возможного нервного субстрата энграммы. В частности, открытие структурных изменений нейронов в ходе формирования памяти дало основание предполагать ключевую роль синтеза белка в ее консолидации, т.е. в переводе информации из кратковременной в долговременную форму ее хранения.
К отличительным особенностям генома нервных клеток относится их высокий по сравнению с другими тканями уровень функционально активных (транскрибируемых) последовательностей ДНК. Характерным для нервных клеток является также прогрессирующее увеличение в них (в течение индивидуальной жизни) числа открытых для синтеза уникальных кодов ДНК, чего не происходит в тканях других органов. В частности, у эмбриона человека в возрасте 22 недель число активных генов в нервной клетке составляет 8,2 %, у взрослого человека эта величина достигает 24,6 %, а в некоторых зонах мозга – 38%, тогда как в мышцах с возрастом она не меняется. Транскрибируемость ДНК и ее синтез в нейронах увеличиваются при обучении животных и содержании их в условиях информационно обогащенной среды.
Известна молекулярная гипотеза памяти П.К. Анохина (1968), согласно которой биохимические процессы, протекающие на уровне протоплазмы, формируют динамические изменения генома нейрона, вызывая перестройку кода РНК, следствием чего является синтез адекватных для данной ситуации новых молекул белка, являющихся хранителем полученной информации. Таким образом, РНК необходима для обучения вообще.
Несмотря на то что механизмы долговременной памяти изучены недостаточно, гипотезы, акцент в которых делается на изменениях белкового обмена нейрона в качестве обязательного условия запоминания (консолидации памяти), в целом подтверждаются. В частности, активно разрабатывается гипотеза X. Хидена о белковой природе памяти: процесс фиксации информации в нервной клетке находит отражение в синтезе белка, в молекулу которого вводится соответствующий следовой отпечаток изменений молекул РНК. При этом молекула белка становится чувствительной к специфическому узору импульсного потока, т.е. она узнает афферентный поток импульсов. Участие нейроспецифических белков в формировании следов памяти доказывается усилением их новообразования в процессе обучения и регистрации расстройства памяти при блокаде их синтеза. Усиление синтеза белков доказывается увеличением внедрения в них метки после предварительного введения меченых аминокислот. Феномен синтеза нейроспецифических белков проявляется в пределах 1 ч обучения и достигает максимума на 3-й и 6-й час (И. П. Ашмарин). Гипотеза о белковой природе долговременной памяти подтверждается многими экспериментами.
Так, при угнетении механизмов, регулирующих синтез нейроспецифических белков, выработанные условные рефлексы при простых формах обучения сохраняются лишь на протяжении нескольких минут, иногда нескольких десятков минут. Затем в динамике этих условных рефлексов отмечаются явные расстройства, указывающие на нарушение процессов сохранения приобретенного навыка. Получены данные о несомненном участии в механизмах памяти двух нейроспецифических белков – S-100 и 14-3-2, содержание которых при обучении увеличивается, особенно в гиппокампе (белок S-100) и в коре головного мозга (белок 14-3-2). Выявлено, что функции памяти связаны с определенными белковыми молекулами. При обучении отмечен синтез специфических полипептидов и мозгоспецифических белков.
Синтез белка ведет к формированию энграммы долговременной памяти. Белок S-100 активно взаимодействует с наружной мембраной и сократительными мембранами нейрона при участии ионов кальция. Белок 14-3-2 – энзим, участвующий в реакциях гликолиза в нейронах. Некоторые авторы рассматривают белок S-100 как глиальный белок.
Если синтез белка, обычно начинающийся в мозгу животного во время сеанса выработки рефлекса и длящийся много часов, блокировать, долговременного научения не произойдет.
Несколько сотен работ, опубликованных на эту тему, дают достаточно ясную картину: синтез белка в мозгу можно временно подавить на 80 – 90% без каких-либо грубых психических нарушений, если не считать памяти. Наиболее выраженная амнезия наблюдается и в случае подавления синтеза белка незадолго до сеанса обучения (и во время него уже не происходит). При этом даже спустя несколько недель повторения опытов усвоение остается неполным. Само по себе выполнение задания, а также ранее заученный материал при угнетении синтеза белка не страдают. Из этого следует, что он необходим лишь на критической стадии консолидации памяти непосредственно во время обучения или сразу после него. Для долговременного хранения информации синтез белка уже не нужен.
Подавление синтеза белка не влияет (по крайней мере, в опытах на животных) на кратковременную память. Это еще один важный аргумент, подтверждающий, что механизмы кратковременной и долговременной памяти различны.
Однако процессы перевода информации из одной формы в другую остаются невыясненными.
Формирование энграммы памяти. В результате активации генома и синтеза специфических белков, в первую очередь в мембранах нервных клеток, в процессе обучения формируется структурно-функциональное объединение нейронов различных структур мозга, представляющее собой энграмму памяти.
Энграмма – это ансамбль нейрональных и глиальных элементов, объединенных синаптическими механизмами. Такое динамическое объединение строится за счет экспрессии геномом определенных белковых молекул (адгезинов или коннектинов), которые встраиваются в специальные области мембран нейронов. Именно эти идентичные по молекулярным свойствам белковые молекулы увеличивают чувствительность нейронов к приему той информации, которая первично вызывала экспрессию этих белков.
Таким образом, изменение белкового метаболизма нейрона, по всей видимости, является решающим звеном в сложнейшей цепи процессов формирования и закрепления следов памяти – энграммы (А. С.Батуев). На основании изложенного долговременную память можно называть структурной памятью (период видимых синтаксических процессов – ультраструктурной продолжительностью около 1 мес, период синтеза белка в клетке – макромолекулярной – месяцы и годы. Эти процессы перекрывают друг друга).
Роль тренировки механизмов памяти. Известно, что у лиц, занимающихся интенсивным умственным трудом, хорошая память сохраняется даже в преклонном возрасте. Косвенным подтверждением этого являются опыты на животных. Так, например, у крыс, воспитанных в обогащенной среде с наличием множества раздражителей и обученных выполнению зрительных задач, кора головного мозга значительно толще, чем у крыс, выращенных в обедненной окружающей среде. В онто- и филогенезе в нервной ткани прогрессивно увеличивается число уникальных кодов ДНК, открытых для синтеза РНК и белков. Транскрибируемость ДНК (а также ее синтез) в нейронах увеличивается как при обучении животных, так и при содержании их в условиях информационно обогащенной среды.
Воспроизведение следов памяти заключается в извлечении информации, сохраняющейся в структурах мозга и, прежде всего, в его генетическом аппарате. Долговременная память в нормальных условиях легко доступна для извлечения.
Процесс вспоминания. В системных процессах извлечения следов памяти ведущая роль принадлежит доминирующей мотивации. На основе предшествующих подкреплений и активации специфических механизмов синтеза, белковых и других биологически активных молекул доминирующая мотивация, формирующаяся в очередной раз на основе соответствующей потребности, распространяясь к геному нейронов, по опережающему принципу активирует в них образование информационных молекул. С помощью этих молекул «оживляется» ранее организованная подкреплением энграмма памяти, которая и определяет процесс воспоминания. При этом молекула белка становится чувствительной к специфическому узору импульсного потока, т.е. она узнает афферентный поток импульсов.
Подсознательная память включается в стереотипные, автоматизированные поведенческие акты. Чаще всего она сопутствует заученным действиям, например исполнению музыкального произведения, спортивного упражнения, вождению автомобиля, чтению стихов и т.д.
Сформированная память может быть вызвана различными ассоциациями: зрительными образами, слуховыми и тактильными воздействиями. Особенно ярки воспоминания, вызванные обонятельными воздействиями. Эмоциональные состояния тоже часто являются причиной соответствующих воспоминаний (Е. А. Громова). Эмоциональная память надмодальна: она воспроизводится разными сенсорными, биологическими или социально значимыми раздражителями. Как правило, эмоциональная память формируется очень быстро, часто с первого раза.
Структурные основы воспоминания. Канадский нейрохирург У. Пенфилд первый показал, что стимуляция медиальных отделов височных долей коры полушарий у пациентов во время нейрохирургических операций вызывала детальные воспоминания событий, имевших место в отдаленном прошлом. При этом новые события оценивались пациентами как уже ранее пережитые.
При электрическом раздражении височных долей возникали живые картины из прошлого в их адекватной временной последовательности – занятия в школе или просмотренный кинофильм. Эти картины были очень ярки – больные переживали все подробности ранее происходивших событий, детали этих событий и соответствующие эмоциональные ощущения. Отмеченные картины четко воспроизводились при повторных стимуляциях медиальных отделов височной доли.
Вместе с тем несмотря на приведенные примеры, демонстрирующие роль височной коры в извлечении опыта из памяти, следует иметь в виду, что память обусловлена деятельностью большого количества объединенных в структурно-функциональные ансамбли нейронов мозга, расположенных как в коре, особенно в лимбических структурах мозга, так и в подкорковых образованиях. Стимуляция височной коры только активирует эту корково-подкорковую энграмму памяти.
Процессы сохранения памяти в значительной степени связаны с функциями гиппокампа, амигдалы и височной коры.
В хранении автоматизированных двигательных навыков существенная роль принадлежит структурам мозжечка.
Забывание. Процесс забывания характеризуется определенной скоростью. Как показали наблюдения над больными и эксперименты на животных, забывание связано с деятельностью структур гиппокампа и височной доли больших полушарий. Больные, у которых повреждены гиппокамп и височная доля, забывают приобретенные навыки очень быстро. У обезьян с разрушенным таламусом не отмечалось такого быстрого забывания, как после удаления гиппокампа. Быстрое забывание связано, по-видимому, с нарушением процесса консолидации памяти, т.е. переводом ее из кратковременной в долговременную память. Ангиотензин-II препятствует процессам забывания условно-рефлекторных оборонительных навыков у крыс.
Дата добавления: 2015-10-16; просмотров: 124 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Кратковременная и промежуточная память | | | Роль отдельных структур мозга в формировании памяти |