Читайте также:
|
БИОЛОГИЯ ЗВЕРЕЙ И ПТИЦ
Методические указания
к проведению лабораторных занятий
Красноярск 2013
Министерство образования и науки Российской Федерации
ФГБОУ ВПО «Сибирский государственный технологический университет»
Кафедра экологии и защиты леса
А. Н. Зырянов, В. А. Тюрин, А. В. Пушкарёв
БИОЛОГИЯ ЗВЕРЕЙ И ПТИЦ
Методические указания к проведению лабораторных занятий
Раздел 2. К ласс млекопитающие
Часть 2. Отряды Насекомоядные, Рукокрылые, Зайцеобразные и Грызуны
для студентов направлений 250100.62 «Лесное дело», профиль подготовки «Лесное хозяйство», 250700.62 «Ландшафтная архитектура», профиль подготовки «Ландшафтное строительство»
очной и заочной форм обучения
Красноярск 2013
Зырянов, А. Н. Биология зверей и птиц: Методические указания к проведению лабораторных занятий по изучению раздела 2 «Биология зверей (класса млекопитающих)». Отряды насекомоядные, рукокрылые, зайцеобразные, грызуны для студентов направлений 250100.62 «Лесное дело», профиль подготовки «Лесное хозяйство», 250700.62 «Ландшафтная архитектура», профиль подготовки «Ландшафтное строительство» очной и заочной форм обучения / А. Н. Зырянов, В. А. Тюрин, А. В. Пушкарёв. — Красноярск: СибГТУ, 2013. — 80 с.
Рецензент: канд. биол. наук, доц. О. П. Ковылина
© ФГБОУ ВПО «Сибирский государственный технологический университет», 2013
Содержание
| Введение | |
| 1. Теоретическая часть | |
| 1.1 Характеристика млекопитающих | |
| 1.2 Способы отлова мелких млекопитающих | |
| 1.3. Систематический указатель видов | |
| 2. Краткие сведения по биоэкологии мелких млекопитающих | |
| 2.1 ОТРЯД НАСЕКОМОЯДНЫЕ - INSECTIVORA BOWDIGH, 1821 | |
| 2.1.1 Ушастый еж - Erinaceus auritus Gmelin, 1770 | |
| 2.1.2 Сибирский крот - Talpa altaica Nikolsky, 1883 | |
| 2.2 Семейство бурозубок - Sorex | |
| 2.2.1 Средняя бурозубка - Sorex caecutiens Laxmann, 1788 | |
| 2.2.2 Крошечная бурозубка - Sorex minutissimus Zimmermann, 1780 | |
| 2.2.3 Обыкновенная бурозубка - Sorex araneus Linnaeus, 1758 | |
| 2.2.4 Арктическая бурозубка - Sorex arcticus Kerr | |
| 2.2.5 Равнозубая бурозубка - Sorex isodon Turov, 1924 | |
| 2.2.6 Малая бурозубка - Sorex minutus L, 1766 | |
| 2.2.7 Бурая бурозубка - Sorex roboratus | |
| 2.2.8 Водяная кутора - Neomys fodiens Pennant, 1771 | |
| 2.3. ОТРЯД РУКОКРЫЛЫЕ - CHIROPTERA LUMENBACH, 1779 | |
| 2.3.1. Ночница восточная (ранее водяная) - Мyotis petax Hollister, 1912 | |
| 2.3.2 Ночница Брандта - Myotis brandtii Eversmann, 1845 | |
| 2.3.3 Ночница прудовая - Myotis dasycneme Bole, 1825 | |
| 2.3.4 Ночница Иконникова - Myotis ikonnikovi Ognev, 1911 | |
| 2.3.5 Ушан Огнева (ранее бурый) - Plecotus ognevi Kishida, 1927 | |
| 2. 3.6 Кожанок северный - Vespertilio nilssonii Keyserling et Blasius, 1839 | |
| 2.3.7 Трубконос большой - Мurina hilgendorfi Peters, 1880 | |
| 2.4. ОТРЯД ЗАЙЦЕОБРАЗНЫЕ - LAGOMORPHA BRANDT, 1855 | |
| 2.4.1Северная (алтайская) пищуха - Ochotona alpine hyperborea Pallas, 1811 | |
| 2.4.2 Даурская пищуха - Ochotona daurica Pallas, 1776 | |
| 2.4.3 Заяц-русак - Lepus europaeus Pallas, 1778 | |
| 2.4.4 Заяц-беляк - Lepus timidus Linnaeus, 1758 | |
| 2.4.5 Заяц-толай - Lepus tolai Pallas, 1778 | |
| 2.5. ОТРЯД ГРЫЗУНЫ - RODENTIA BOWDICH, 1821 | |
| 2.5.1 Обыкновенная белка - Sciurus vulgaris Linnaeus, 1758 | |
| 2.5.2 Азиатский бурундук - Tamias sibiricus Laxmann, 1769 | |
| 2.5.3 Тарбаган - Marmota sibirica Radde, 1862 | |
| 2.5.4 Длиннохвостый суслик - Spermophilus undulatus Pallas, 1779 | |
| 2.5.5 Летяга - Pteromys volans Linnaeus, 1758 | |
| 2.5.6 Бобр - Castor fiber Linnaeus, 1758 | |
| 2.5.7 Ондатра - Ondatra zibethica Linnaeus, 1766 | |
| 2.5.8 Красно-серая полевка - Clethrionomys rufocanus Sundevall, 1846-1847 | |
| 2.5.9 Красная полевка - Clethrionomys rutilus Pallas, 1779 | |
| 2.5.10 Полевка-экономка - Microtus oeconomus Pallas, 1776 | |
| 2.5.11 Узкочерепная полевка - Microrus gregalia Pallas, 1779 | |
| 2.5.12 Темная, или пашенная полевка - Microtus agrestis L., 1761 | |
| 2.5.13 Лесной лемминг - Myopus schisticolor Lilljeborg, 1844 | |
| 2.5.14 Водяная полевка - Arvicola terrestris L., 1758 | |
| 2.5.15 Полевая мышь - Apodeinus agrarius Pallas,1771 | |
| 2.5.16 Восточноазиатская (азиатская) мышь - Apodemus penninsulae Thomas, 1906 | |
| 2.5.17 Лесная мышь - Apodemus uralensis L., 1758 | |
| 2.5.18 Домовая мышь - Mus musculus L., 1758 | |
| 2.5.19 Серая крыса - Rattus norvegicus Berkenhout, 1769 | |
| 2.5.20 Лесная мышовка - Sicista betulina Pall.,1775 | |
| 3. Определение следов жизнедеятельности | |
| 3.1.Экскременты | |
| 3.2. Корм | |
| 3.3. Следы от погрызов | |
| 3.4. Тропы, убежища | |
| 3.5. Звуки | |
| 4. Практическая часть | |
| Лабораторная работа № 1 Морфология и анатомия млекопитающих (6 часов) | |
| Лабораторная работа № 2 Определение семейств, родов и видов отрядов насекомоядные, рукокрылые, зайцеобразные, грызуны (8 часов) | |
| 5. Библиографический список | |
| Приложение А | |
| Приложение Б |
Введение
Высокое видовое разнообразие и численность позвоночных в лесу обусловлена сочетанием сред обитания на индивидуальных участках (почвенная, водная, воздушная, древесная), что обеспечивает широкий выбор гнездовых, комовых и защитных условий в разные сезоны.
Лесные позвоночные, несмотря на незначительную долю биомассы (до 1 %), являются одним из важнейших компонентов леса, функционально связанного с растительностью и другими организмами, образую консорции (плеяды) древесных пород. Они завершают энергетическую пирамиду лесной экосистемы и выполняют ресурсную, эстетическую и лесообразующую функцию.
Позвоночные регулируют вегетативный рост древесных пород, особенно на стадии лесовозобновления, уничтожают и участвуют в распространении семян, минерализуя почву облегчают их прорастание, питаясь фитофагами регулируют численность вредителей, разрушая мертвую древесину ускоряют процесс деструкции органической массы, продукты экскреции усиливают почвенные процессы.
Подготовленный лабораторный практикум представляет собой набор повидовых очерков наиболее распространенных и тестовых видов мелких млекопитающих, необходимых для проведения лабораторных занятий по дисциплинам «Биология зверей и птиц» и «Биоэкология позвоночных». Лабораторный практикум «Биоэкология мелких млекопитающих» является второй частью курса дисциплин «Биология зверей и птиц» и «Биоэкология позвоночных». В него входит раздел Биология зверей (класса млекопитающих) и две темы:
Тема 1. Общая характеристика класса млекопитающих.
Тема 2. Отряды насекомоядные, рукокрылые, зайцеобразные, грызуны.
В каждом очерке представлено: описание вида, его распространение, экология, хозяйственное значение.
Роль и место лабораторного практикума. Выполнение данного курса лабораторных работ тесно связано с такими дисциплинами, как экология, охотоведение, лесоведение и лесоводство, лесные культуры, лесоустройство.
Цель и задачи лабораторного практикума. В процессе выполнения лабораторных занятий студент должен сформировать представление об экологических требованиях мелких млекопитающих. Для этого приобретаются навыки определения, выясняются характерные морфологические особенности. Из описания вида выбираются экологические особенности по питанию, образу жизни, распространению, местообитаниям, размножению, динамике численности.
Студенты знакомятся с характеристикой класса млекопитающие по схеме: систематика, особенности анатомического, морфологического и физиологического строения, размножения, питания, экологии. Курс лабораторных занятий предусматривает изучение хозяйственной и биоценотической роли зверей в лесу, воздействие позвоночных на лесообразовательный процесс (семена, вегетативные части, вредителей леса). Анализируются экологические условия, провоцирующие вредную деятельность позвоночных.
Форма проведения занятий. Знакомство с текстом практикума, определение представленного биоматериала, конспектирование и зарисовка, заполнение таблиц и схем.
В результате выполнения лабораторных работ студент должен знать экологические особенности видов и экологических групп, их оптимальные биотопы, приемы отлова и определения, а также уметь использовать полученные знания на практике.
А) по направлению 250100.62 Лесное дело
ОК 1: владение культурой мышления, способность к обобщению, анализу, восприятию информации, постановке цели и выбору путей ее достижения;
ОК 3: готовность к кооперации с коллегами и работе в коллективе;
ОК 4: способность находить организационно-управленческие решения в нестандартных ситуациях и готов нести за них ответственность;
ОК 7: осознание социальной значимости своей будущей профессии, обладанием высокой мотивации к выполнению профессиональной деятельности.
ПК 1: использование основных законов естественнонаучных дисциплин в профессиональной деятельности, умение применять методы математического анализа и моделирования, экспериментального исследования;
ПК 2: владение основными методами, способами и средствами получения, хранения, переработки информации, навыками работы с компьютером, как средством управления информацией;
ПК 5: знание роли основных компонентов лесных и урбо – экосистем: растительного и животного мира, почв, поверхностных и подземных вод, воздушных масс тропосферы в формировании устойчивых, высокопродуктивных лесов;
ПК 37: способность применять современные методы исследования объектов растительного и животного мира;
Б) по направлению 250700.62 Ландшафтная архитектура
ОК 1: владение культурой мышления, способность к обобщению, анализу, восприятию информации, постановке цели и выбору путей её достижения;
ОК 2: умение логически верно, аргументировано и ясно строить устную и письменную речь;
ПК 1: способность использовать основные законы естественнонаучных дисциплин в профессиональной деятельности, применять методы математического анализа и моделирования, экспериментального исследования;
ПК 5: понимание роли основных компонентов урбо- экосистем: растительного и животного мира, почв, поверхностных и подземных вод, воздушных масс тропосферы в формировании устойчивых, жизнестойких, эстетически выразительных зелёных насаждений, сооружений, оборудования объектов ландшафтной архитектуры;
ПК 17: умение в полевых условиях при проведении ландшафтного анализа определять систематическую принадлежность, названия основных видов вредных и полезных лесных насекомых и других хозяйственно значимых организмов.
Лабораторный практикум предназначен для изучения курса «Биология зверей и птиц» общим объемом 108 часов. Изучение курса завершается зачётом по теоретической части с предварительным допуском по определению позвоночных, а также следов их жизнедеятельности.
1. Теоретическая часть
1.1 Характеристика млекопитающих
Некоторые части скелета млекопитающих, особенно череп, устроены проще, чем у их предков-пресмыкающихся. Например, каждая ветвь (правая и левая) их нижней челюсти состоит из одной кости, а у рептилий – из нескольких. У зверей верхняя челюсть (межчелюстная кость спереди и верхнечелюстная сзади) полностью слита с черепной коробкой, тогда как у некоторых рептилий она соединена с ней подвижно эластичными связками. У млекопитающих верхние зубы находятся только на межчелюстной и верхнечелюстной костях, а у примитивных позвоночных они могут быть и на других костных элементах крыши ротовой полости, в том числе на сошниках (возле носовых ходов) и небных костях (рядом с верхнечелюстными). У млекопитающих обычно две пары функциональных конечностей, но у некоторых водных форм, например китов (Cetacea) и сирен (Sirenia), сохранились только передние. Все звери теплокровны и дышат атмосферным воздухом. От всех остальных позвоночных, за исключением птиц и крокодилов, они отличаются четырехкамерным сердцем и полным разделением в ней артериальной и венозной крови. Однако, в отличие от птиц и крокодилов, зрелые красные кровяные клетки (эритроциты) млекопитающих лишены ядер. За исключением наиболее примитивных представителей класса, все млекопитающие живородящие и кормят детенышей молоком, вырабатываемым молочными железами матери. Первозвери, или однопроходные, например утконос, откладывают яйца, но вылупившиеся из них детеныши также питаются молоком. У некоторых видов они рождаются хотя и полностью сформированными, но голыми (без шерсти) и беспомощными, а глаза у них в течение некоторого времени остаются закрытыми. У других зверей, особенно копытных (козы, лошади, олени и т.д.) детеныши появляются на свет полностью одетыми шерстью, с открытыми глазами и почти сразу способны стоять и передвигаться. У сумчатых, например кенгуру, детеныши рождаются недоразвитыми и донашиваются в течение некоторого времени в кармане на животе матери.
Шерсть. Наличие покрывающей тело шерсти – отличительный признак зверей: только у них образуются волосы, т. е. нитевидные ороговевшие выросты кожи (эпидермиса). Основная функция шерстного покрова – теплоизоляция организма, облегчающая терморегуляцию, однако он служит и для многих других целей, в частности защищает кожу от повреждений, может маскировать животное за счет своей окраски или конфигурации или демонстрировать его половую принадлежность. Волосяной покров характерен почти для всех зверей, кроме китообразных, ящеров и броненосцев. На ранних стадиях эмбрионального развития млекопитающих волосы в первоначальном виде напоминают зачатки чешуи, что дает основание рассматривать их как результат эволюционного преобразования чешуйчатого покрова пресмыкающихся - предков млекопитающих. У некоторых зверей волосы частично видоизменились в жесткую щетину (кабаны, образовали пучки твердых волос - вибриссы (на морде у кошек) или превратились в иглы, как, например, у дикобраза и ежа. Все преобразования волосяного покрова связаны с приспособлением тех или иных видов к выполнению определенных биологических функций. Мех у зверей разных видов различен. У одних, например, кротов, некоторых землероек - только короткий, с густым подшерстком, у других - многих хищников - кроме густого подшерстка хорошо развиты и многочисленные остевые волосы, у третьих - волосяной покров почти или полностью состоит из остевых (кабаны, тюлени, олени). У некоторых млекопитающих остевые волосы местами бывают очень длинными и образуют гривы, хвосты и т. д. Отдельные волоски в большинстве случаев цилиндрические или овальные в сечении, хотя у некоторых видов они практически плоские. Микроскопическое изучение показывает, что ствол волоса (над кожей и непосредственно под ней) – это компактный гибкий стержень, состоящий из отвердевших мертвых клеток. Типичный ствол состоит из трех концентрических слоев: центральной губчатой сердцевины, образованной рыхло лежащими прямоугольными клетками, часто с небольшими прослойками воздуха между ними, среднего коркового слоя, составляющего основную часть волоса и образованного продольно расположенными вплотную друг к другу веретеновидными клетками, и тонкой внешней кожицы (кутикулы) из чешуевидных, накладывающихся друг на друга клеток, свободные края которых направлены к свободному концу волоса. Клетки волоса формируются под кожей внутри волосяного мешочка (фолликула) и выталкиваются наружу новыми клетками, образующимися под ними. По мере удаления от корня, т.е. источника питания, клетки отмирают и обогащаются кератином – нерастворимым белком в виде длинных тонких волокон. Волокна кератина химически связываются между собой, что придает волосу прочность. Цвет волос зависит от нескольких факторов. Наличие пигментов (окрашивающих веществ), называемых меланинами. Несмотря на то, что название этих пигментов происходит от слова «черный», их цвет варьирует от желтого до красного, коричневого и черного. Меланины могут появляться в отдельных клетках волоса по мере их роста и удаления от фолликула. Наличие или отсутствие меланина, его цвет и количество, а также доля прослоек воздуха между клетками ствола в совокупности определяют все разнообразие окрасок волоса. В принципе, можно сказать, что его цвет зависит от поглощения и отражения света меланином (главным образом коркового слоя) и его рассеивания стенками воздушных прослоек сердцевины. Например, черный волос содержит оптически плотный очень темный меланин и в коре и в сердцевине, поэтому отражает лишь очень небольшую часть световых лучей. И напротив, шерсть белого медведя вообще лишена пигмента, а ее цвет определяется равномерным рассеиванием света. Разнообразие строения волос в первую очередь связано с формой кутикулярных клеток и расположением сердцевинных. Конкретные виды животных, как правило, характеризуются определенной структурой шерсти, поэтому с помощью микроскопа обычно можно определить ее таксономическую природу. Изменение окраски и характера волосяного покрова в результате линьки называется сезонной изменчивостью, которая широко распространена у наземных млекопитающих. У одних наблюдается осветление меха зимой и осенью, иногда до чисто белого, как у зайца-беляка, горностая, ласки, копытного лемминга. У многих пушных зверей, наоборот, появляется более яркая и контрастная окраска. Волосяной покров животных зимой всегда длиннее и пышнее, хорошо развита подпушь. Кроме сезонной смены меха существует возрастная линька, при которой ювенальный наряд молодых животных сменяется дефинитивным взрослых.
Встречаются также особые отклонения в окраске, выражающиеся в появлении черных или желтых пятен (частичный альбинизм, меланизм и хромизм. Реже в природе встречаются абсолютно белые, черные или желтые особи. Полные альбиносы обычно отторгаются нормально окрашенными особями того же вида и долго не могут существовать в естественных условиях. Наиболее широко распространено явление альбинизма и меланизма среди хомяков, кротов и землероек.Зубы, присутствующие у подавляющего большинства млекопитающих, представляют собой твердые структуры, которые развиваются из особых соединительнотканных (мезодермальных) клеток – одонтобластов и состоят в основном из фосфата кальция (апатита), т.е. по химическому составу очень похожи на кости. Однако фосфат кальция кристаллизуется и сочетается с другими веществами по-разному, так что в результате образуются различные зубные ткани – дентин, эмаль и цемент. В основном зуб состоит из дентина. (Бивни слона и соответственно слоновая кость – сплошной дентин; небольшое количество эмали, покрывающее сначала конец бивня, быстро стирается.) Полость в центре зуба содержит питающую его «пульпу» из мягкой соединительной ткани, кровеносных сосудов и нервов. Обычно выступающая наружу поверхность зуба хотя бы частично покрыта тонким, но исключительно твердым слоем эмали (самое твердое вещество в организме), которая образуется особыми клетками – амелобластами (адамантобластами). Зуб крепится в ячейке на челюсти с помощью цемента, который по твердости занимает промежуточное положение между эмалью и дентином. Он может также присутствовать внутри самого зуба и на его жевательной поверхности, например у лошадей.
Зубы млекопитающих обычно делятся на четыре группы в соответствии со своими функциями и местоположением: резцы, клыки, премоляры (малые коренные, ложнокоренные, или предкоренные) и моляры (коренные). Резцы находятся в передней части рта (на межчелюстных костях верхней челюсти и, как и все зубы нижней челюсти, на зубных костях). У них режущие края и простые конические корни. Они служат в основном для удерживания пищи и откусывания ее частей. Клыки (у кого они есть) представляют собой обычно длинные заостренные на конце стержни. Их, как правило, четыре (по 2 верхних и нижних), и расположены они позади резцов: верхние – в передней части верхнечелюстных костей. Клыки используются в основном для нанесения проникающих ран при нападении и защите, удерживания и переноса пищи. Премоляры находятся между клыками и молярами. У некоторых примитивных млекопитающих их по четыре с каждой стороны верхней и нижней челюсти (всего 16), но большинство групп в ходе эволюции утратили часть ложнокоренных зубов, и у человека, например, их всего 8. Моляры, расположенные в задней части челюстей, вместе с премолярами объединяются в группу щечных зубов. Ее элементы могут различаться по размерам и форме в зависимости от характера питания вида, но обычно обладают широкой, ребристой или бугорчатой жевательной поверхностью для раздавливания и перемалывания пищи.
Некоторые млекопитающие, в частности ленивцы, зубатые киты и утконос, развивают всего одну смену зубов на протяжении всей жизни (у утконоса она присутствует только на стадии эмбриона) и называются монофиодонтными. Однако большинство зверей – дифиодонтные, т.е. у них две смены зубов – первая, временная, называемая молочной, и постоянная, свойственная взрослым животным. У них резцы, клыки и премоляры раз в жизни полностью замещаются, а моляры вырастают без молочных предшественников, т.е. по сути дела являются поздно развивающейся частью первой смены зубов. Сумчатые занимают промежуточное положение между монофиодонтами и дифиодонтами, поскольку сохраняют все молочные зубы, кроме сменяющегося четвертого премоляра. (У многих из них ему соответствует третий щечный зуб, поскольку один премоляр утрачен в ходе эволюции.) Поскольку у разных видов млекопитающих зубы гомологичны, т.е. одинаковы по эволюционному происхождению (за редкими исключениями, например, у речных дельфинов зубов больше сотни), каждый из них занимает строго определенное положение относительно других и может быть обозначен порядковым номером. В результате характерный для вида зубной набор нетрудно записать в виде формулы.
Поскольку млекопитающие – двусторонне-симметричные животные, такую формулу составляют только для одной стороны верхней и нижней челюстей, помня, что для подсчета общего числа зубов необходимо умножить соответствующие цифры на два.
Развернутая формула (I – резцы, C – клыки, P – премоляры и M – моляры, верхняя и нижняя челюсти – числитель и знаменатель дроби) для примитивного набора из шести резцов, двух клыков, восьми ложнокоренных и шести коренных зубов выглядит следующим образом:
 (х2 = 44, полное число зубов).
(х2 = 44, полное число зубов).
Однако обычно используется сокращенная формула, где указано лишь общее число зубов каждого типа. Для вышеприведенного примитивного зубного набора она выглядит так:

Для домашней коровы, у которой нет верхних резцов и клыков, запись принимает следующую форму:

а у человека выглядит так:
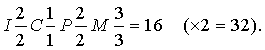
Поскольку все типы зубов расположены в неизменном порядке – I, C, P, M, – зубные формулы часто еще более упрощают, опуская эти буквы. Тогда для человека получаем:
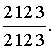
Некоторые зубы, выполняющие особые функции в ходе эволюции, могут претерпевать очень сильные изменения. Например, в отряде хищных (Carnivora), т.е. у кошек, собак и т.п., четвертый верхний премоляр (обозначаемый P4) и первый нижний моляр (M1) крупнее всех прочих щечных зубов и снабжены острыми, как лезвия, режущими кромками. Эти зубы, называемые хищническими, расположены друг напротив друга и действуют наподобие ножниц, разрезая мясо на куски, которые животному удобнее проглотить. Система P4/M1 – отличительный признак отряда Carnivora, хотя ее функцию могут выполнять и другие зубы. Например, молочный набор Carnivora не содержит моляров, и в качестве хищнических используются только премоляры (dP3/dP4).
Скелет. У млекопитающих, как и у всех позвоночных, скелет состоит из большого числа костей, развивающихся независимо и соединенных между собой связками и соединительной тканью. У некоторых видов он глубоко специализирован, но принцип его строения одинаков у всех представителей класса. Это фундаментальное сходство хорошо заметно при сравнении крайних вариантов, например дельфинов с практически отсутствующей шеей, позвонки которой толщиной с бумагу, и жирафов с таким же числом, но сильно вытянутых шейных позвонков.
Череп млекопитающих сочленяется с позвоночным столбом двумя округлыми костными выступами в своей задней части – затылочными мыщелками. Для сравнения, у черепа рептилий только один затылочный мыщелок, т.е. только одна точка сочленения с позвоночником.
Первые два позвонка называются атлантом и эпистрофеем. Вместе со следующими пятью они составляют семь шейных позвонков. Это число типично для всех млекопитающих, кроме ленивцев (от шести до девяти). Затем идет самый крупный, грудной отдел позвоночника; к его позвонкам прикреплены ребра. Далее следуют поясничные (между грудной клеткой и тазом) и крестцовые позвонки. Последние срослись между собой и сочленены с тазовыми костями. Число хвостовых позвонков сильно варьирует в зависимости от вида животного и доходит до нескольких десятков.
У разных млекопитающих количество ребер, окружающих многие жизненно важные органы, неодинаково. Обычно они плоские и изогнуты дугой. Каждое ребро подвижно сочленено одним концом (проксимальным) со спинным позвонком, а другим концом (дистальным) передние ребра (у человека – верхние) прикрепляются к грудине с помощью хряща. Их называют истинными в отличие от задних (у человека – нижних), не соединенных с грудиной и называемых ложными. Дистальный конец этих ребер либо прикреплен к хрящевой части последнего истинного ребра, либо остается свободным, и тогда их называют колеблющимися.
Грудина состоит из ряда слитых вместе более или менее уплощенных костей и с каждой стороны соединяется хрящом с ребрами. У летучих мышей она несет выступающий киль для прикрепления мощных летательных мышц. Похожий киль на грудине есть у летающих птиц и пингвинов (которые «летают» под водой), тогда как нелетающие птицы вроде страуса его лишены.
Лопатка – это широкая плоская кость со срединным гребнем (остью) на наружной поверхности. Ключица одним концом соединяется с верхним краем грудины, а другим – с плечевым отростком (акромионом) ости лопатки. Ключица укрепляет плечо, поэтому она свойственна прежде всего тем млекопитающим (например, приматам), которые интенсивно используют передние конечности для хватания. Она также присутствует у примитивных видов, в частности однопроходных, поскольку является частью предкового (рептильного) плечевого пояса, скелетного образования, связывающего переднюю конечность с осью тела. Ключица была редуцирована или утрачена в ходе эволюции таких групп млекопитающих, которым она не нужна. Например, она рудиментарна у лошади, поскольку лишь мешала бы удлинению ее шага (осталась только маленькая полоска, окруженная мышцами), и отсутствует у китов. Таз (тазовый пояс) служит для прикрепления задних конечностей к позвоночнику.
Конечности. Самая верхняя кость передней конечности (руки у человека) – плечевая. Она причленяется к лопатке с помощью шаровидного сустава, а нижним концом связана с двумя костями предплечья (подплечья) – лучевой и локтевой. Запястье обычно состоит из шести-восьми небольших косточек (у человека их восемь), которые соединяются с костями пясти, образующими «ладонь» кисти. Кости пальцев называются фалангами. Бедренная кость задней конечности (ноги у человека) сочленена шаровидным суставом с тазом. Скелет голени состоит из двух костей – большой и малой берцовых. Затем идет стопа, т.е. предплюсна из нескольких костей (у человека – семь), соединяющихся с костями плюсны, к которым причленены фаланги пальцев.
Число пальцев стоп и кистей зависит от вида млекопитающего – от одного до пяти. Пять – примитивное (предковое) состояние, а, например, у лошади, относящейся к эволюционно продвинутым формам, всего по одному пальцу и на передних и на задних конечностях (анатомически это сильно увеличенный средний, т.е. третий, палец, а остальные утрачены в ходе специализации). У оленя функциональны крупные третий и четвертый пальцы, образующие раздвоенное копыто; второй и пятый маленькие, не достающие до земли, а первый («большой») отсутствует.
У большинства млекопитающих концы пальцев защищены когтями, ногтями или копытами, представляющими собой ороговевшие производные эпидермиса (наружного слоя кожи). Внешний вид и функции этих структур сильно различаются, но их общий план строения один и тот же.
Млекопитающие, опирающиеся при ходьбе на всю подошву, т.е. на пясть и плюсну, как, например, медведи и люди, называются стопоходящими, передвигающиеся с опорой только на пальцы (например, кошки и собаки) – пальцеходящими, а копытные формы (корова, лошадь, олень) – фалангоходящими.
Полость тела всех зверей разделена надвое мускулистой перегородкой, называемой диафрагмой. Спереди (у человека – сверху) находится грудная полость, вмещающая легкие и сердце, а сзади (у человека – снизу) – брюшная полость с остальными внутренними органами, кроме почек. Диафрагма есть только у млекопитающих: она участвует в вентиляции легких.
Сердце млекопитающих разделено на четыре камеры – два предсердия и два желудочка. Каждое предсердие сообщается с желудочком той же стороны тела, но это отверстие снабжено клапаном, позволяющим крови двигаться только в одном направлении. Обедненная кислородом кровь, возвращаясь в сердце от органов тела, поступает в правое предсердие через крупные вены, называемые полыми. Затем она проталкивается в правый желудочек, который перекачивает ее к легким через легочные артерии. В легких кровь насыщается кислородом и отдает углекислый газ. Богатая кислородом кровь поступает затем в легочные вены, а из них – в левое предсердие. Потом она проталкивается из него в левый желудочек, который перекачивает ее через самую крупную артерию – аорту – ко всем органам тела.
Легкие – это губчатая масса, состоящая из многочисленных заполненных воздухом проходов и камер, окруженных сетью капилляров. Проходя через эту сеть, кровь поглощает кислород из закачиваемого в легкие воздуха и одновременно выделяет в него углекислый газ.
Нормальная температура крови у разных видов млекопитающих неодинакова, а у многих летучих мышей, грызунов и ряда других видов заметно падает во время сна и сезонной спячки. Обычно близкая к 38 °C, в последнем случае она может приближаться к точке замерзания.
Характерная для млекопитающих «теплокровность», т.е. способность поддерживать постоянную температуру тела, – понятие относительное. Обитающие в пустыне животные каждый день подвергаются действию сильной жары, что также сказывается на температуре их тела; у верблюдов, например, она может изменяться в течение суток почти на 6○С.
Желудок большинства млекопитающих состоит из одного отдела, однако у некоторых видов их несколько, например, четыре у жвачных, т.е. парнокопытных животных типа коров, оленей и жирафов, пережевывающих жвачку. Верблюдов называют «ложножвачными», поскольку они, хотя и жуют жвачку, отличаются от «истинных» жвачных трехкамерным желудком и некоторыми признаками зубов, ног и других органов.
Нижний конец желудка открывается в тонкую кишку, которая, в свою очередь, ведет в толстую, а та – в прямую. На границе тонкой и толстой кишок от пищеварительного тракта ответвляется слепая кишка. У человека и некоторых других животных она оканчивается маленьким рудиментом – червеобразным отростком (аппендиксом). Строение и роль слепой кишки сильно различаются в зависимости от вида животного. Например, у жвачных и лошадей она выполняет важную функцию бродильной камеры для переваривания растительных волокон и исключительно длинна, а у других млекопитающих сравнительно мала, хотя и принимает активное участие в пищеварении.
Молочные железы вырабатывают молоко для питания детенышей. Эти структуры закладываются у представителей обоих полов, но у самцов недоразвиваются. У всех млекопитающих, кроме утконоса и других однопроходных, протоки молочных желез открываются на мясистых выростах – сосках, которые детеныши, питаясь, захватывают ртом. У некоторых видов, например коров, протоки молочной железы сначала впадают в камеру, называемую цистерной, где накапливается молоко, поступающее затем наружу через длинные трубчатые соски. У однопроходных сосков нет, и молочные протоки открываются порообразными отверстиями на коже.
Нервная система функционирует как единое целое с органами чувств, например глазами, и управляется у млекопитающих головным мозгом. Самая крупная часть последнего называется большими полушариями (в затылочной области черепа находятся два более мелких полушария мозжечка). Головной мозг соединяется со спинным. У всех млекопитающих, за исключением однопроходных и сумчатых, в отличие от других позвоночных правое и левое большие полушария соединены между собой компактным пучком нервных волокон, называемым мозолистым телом. В мозгу однопроходных и сумчатых мозолистого тела нет, но соответствующие области полушарий тоже соединяются нервными пучками. Спинной мозг – основной нервный ствол тела – проходит через канал, образованный отверстиями позвонков, и тянется от головного мозга до поясничного или крестцового отдела позвоночника, в зависимости от вида животного. С каждой стороны спинного мозга симметрично отходят нервы к различным частям тела.
Осязание в общих чертах обеспечивается определенными нервными волокнами, бесчисленные окончания которых находятся в коже. Эта система обычно дополняется волосками, действующими, как рычаги, для нажатия на пронизанные нервами участки.
Зрение более или менее развито у всех млекопитающих, хотя у некоторых слепышей маленькие недоразвитые глаза покрыты кожей и вряд ли даже способны отличать свет от тьмы. Животное видит отраженный от объектов свет, поглощаемый глазом, который передает соответствующие сигналы в головной мозг для распознавания. Другими словами, сами глаза не «видят», а действуют только как преобразователи световой энергии.
Одна из проблем получения четкого зрительного образа – преодоление хроматической аберрации, т.е. размытой цветной каймы, появляющейся по краям изображения, сформированного простой линзой (несоставным прозрачным предметом с двумя противоположными поверхностями, из которых по крайней мере одна криволинейна). Хроматическая аберрация является неотъемлемым свойством хрусталика глаза и возникает из-за того, что он, как простая линза, преломляет свет с более короткой длиной волны (например, фиолетовый) сильнее, чем длинноволновые лучи (например, красные). Таким образом, лучи всех длин волн фокусируются не в одной точке, давая четкое изображение, а одни ближе, другие – дальше, и изображение получается расплывчатым. В механической системе типа фотокамеры хроматическую аберрацию корректируют, склеивая линзы с различной взаимокомпенсирующей преломляющей способностью. Глаз млекопитающих решает эту проблему путем «отсекания» большей части коротковолнового света. Желтоватый хрусталик действует как желтый фильтр: он поглощает почти весь ультрафиолет (отчасти поэтому человек его и не воспринимает) и часть сине-фиолетового участка спектра.
Не весь свет, проникающий через зрачок и достигающий светочувствительной сетчатки, используется для зрения. Часть его проходит сквозь сетчатку и поглощается лежащим под ней пигментным слоем. Для ночных животных это означало бы слишком большие потери незначительного количества доступного света, поэтому у многих таких видов дно глаза зеркальное: оно отражает неиспользованный свет обратно на сетчатку для дополнительной стимуляции ее рецепторов. Именно этот отраженный свет заставляет глаза некоторых млекопитающих «светиться» в темноте. Зеркальный слой называется tapetumlucidum (зеркальце). У млекопитающих два основных типа зеркальца. Первый – волокнистый, характерный для копытных. Их зеркальце в основном состоит из блестящего слоя волокон соединительной ткани. Второй тип – клеточный, например у хищных. В этом случае оно состоит из нескольких слоев уплощенных клеток, содержащих волокнообразные кристаллы. Зеркальце обычно расположено в сосудистой оболочке за сетчаткой, но, например, у некоторых летучих мышей и у виргинского опоссума оно погружено в саму сетчатку. Цвет, которым светятся глаза, зависит от количества крови в капиллярах сосудистой оболочки и содержания родопсина (пурпурного светочувствительного пигмента) в палочковидных элементах сетчатки, через которую проходит отраженный свет.
Несмотря на широко распространенное убеждение, что цветовое зрение – редкость у млекопитающих, большинство которых якобы видит только оттенки серого, судя по накапливающимся фактам, многие виды, включая домашних кошек и собак, все же, хотя бы в некоторой степени, различают и цвета. Цветовое зрение, вероятно, наиболее развито у приматов, но известно также у лошади, жирафа, виргинского опоссума, нескольких видов белок и многих других зверей.
Слух хорошо развит у многих млекопитающих, причем для 20% их видов он во многом заменяет зрение. Слуховой аппарат состоит из трех основных частей. Млекопитающие – единственная группа животных с хорошо развитым наружным ухом. Ушная раковина улавливает звуковые волны и направляет их на барабанную перепонку. С внутренней стороны от нее находится следующий отдел – среднее ухо, заполненная воздухом камера с тремя косточками (молоточком, наковальней и стремечком), которые механически передают колебания от барабанной перепонки к внутреннему уху. Оно включает в себя улитку – спирально закрученную, заполненную жидкостью трубку с волосовидными выростами внутри. Звуковые волны вызывают колебания жидкости и, опосредованно, движение волосков, что служит стимуляцией нервных клеток у их основания.
Частотный диапазон воспринимаемых звуков зависит от вида животного. Многие мелкие млекопитающие слышат «ультразвук» с частотами, которые слишком высоки для слуха человека. Ультразвук особенно важен для видов, использующих эхолокацию – улавливание отраженных звуковых волн (эха) для распознавания объектов в окружающей среде. Такой способ ориентации характерен для летучих мышей и зубатых китов. С другой стороны, многие крупные млекопитающие могут улавливать низкочастотный «инфразвук», который человек также не слышит.
Обоняние связано с тонкими чувствительными мембранами (обонятельной слизистой) в задней части носовой полости. Они улавливают молекулы пахучих веществ, присутствующие во вдыхаемом воздухе. Обонятельная слизистая состоит из нервных и опорных клеток, покрытых слоем слизи. Окончания ее нервных клеток несут пучки обонятельных «ресничек» числом до 20, которые в совокупности образуют как бы ворсистый ковер. Реснички служат рецепторами запахов, причем густота их «ковра» зависит от вида животного. У человека, например, их бывает до 20 млн. на площади 5 см2, а у собаки – более 200 млн. Пахучие молекулы растворяются в слизи и попадают в особые чувствительные ямки на ресничках, стимулируя нервные клетки, которые посылают в головной мозг импульсы для анализа и распознавания.
Коммуникация. Звуковая. Млекопитающие используют при общении звуки, издавая, например, сигналы тревоги, угрозы или призыва к спариванию (некоторые животные, в частности определенные виды оленей, подают голос только в сезон размножения). Ряд видов, включая кроликов, обладает хорошо развитыми голосовыми связками, но пользуется ими только при крайнем стрессе. Неголосовая звуковая коммуникация известна у многих млекопитающих: кролики, например, стучат для этого по земле лапой, белоногие хомячки барабанят передними лапами по полым предметам, а самцы оленей трещат рогами по ветвям. Звуковая коммуникация играет важную роль в социальных взаимодействиях зверей, поскольку в общем они могут выразить звуками все основные эмоции.
Летучие мыши издают звуки для эхолокации, позволяющей им ориентироваться в темноте или в мутной воде, где зрения для этого было бы явно недостаточно.
Зрительная. Млекопитающие общаются не только с помощью звуков. Например, у некоторых видов белая нижняя сторона хвоста при необходимости демонстрируется сородичам в качестве зрительного сигнала. «Чулки» и «маски» определенных антилоп также широко используются для демонстраций их состояния. Особый пример зрительной коммуникации наблюдается у американского вилорога, посылающего сообщения другим особям своего вида в радиусе 6,5 км с помощью участка длинной белой шерсти на крупе. Вспугнутый зверь резко распушает эти волосы, которые как бы вспыхивают на солнечном свете, становясь хорошо заметными на большом расстоянии.
Химическая. Запахи, определяющиеся различными химическими веществами в моче, фекалиях и секрете желез, широко используются млекопитающими при социальных взаимодействиях, например для мечения территории или распознавания подходящих для спаривания партнеров. В последнем случае запах позволяет не только отличать самцов от самок, но и определять стадию репродуктивного цикла конкретной особи. Химические сигналы, применяемые для внутривидового общения, называются феромонами (от греч. pherein – нести и hormon – возбуждать, т.е. феромоны «переносят возбуждение» от одной особи к другой). Они делятся на два функциональных типа: сигнальный и мотивирующий. Сигнальные феромоны (релизеры) вызывают специфические поведенческие реакции другого животного, например привлекают особей противоположного пола, заставляют следовать по оставленному пахучему следу, спасаться бегством или нападать на врага. Мотивирующие феромоны (праймеры) приводят к физиологическим изменениям у сородичей. Например, достижение половозрелости.
Размножение. Рыбы и амфибии обычно откладывают яйца (икринки) в воду. Их яйца снабжены мембранами, которые помогают развивающимся эмбрионам избавляться от отходов и поглощать питательные вещества, в первую очередь из богатого калориями желтка. Желточный мешок и другие мембраны такого типа находятся снаружи от эмбриона, поэтому называются внезародышевыми оболочками.
Рептилии стали первыми позвоночными, которые приобрели три дополнительные внезародышевые оболочки, позволяющие откладывать яйца на сушу и обеспечивающие развитие без водного окружения. Эти оболочки дали возможность эмбриону получать питательные вещества, воду и кислород, а также выделять продукты обмена, находясь в неводной среде.
Самая внутренняя из них – амнион – образует мешок, заполненный солоноватой жидкостью. Она окружает зародыш, обеспечивая ему жидкую среду, аналогичную той, в которую погружены эмбрионы рыб и амфибий в воде, а обладающие ею животные называются амниотами. Самая наружная оболочка – хорион – вместе со средней (аллантоисом) выполняет другие важные функции. Оболочка, окружающая икринку рыб, также называется хорионом, но эта структура у них функционально сопоставима с т.н. блестящей оболочкой яйца млекопитающих, присутствующей еще до его оплодотворения.
Звери унаследовали внезародышевые оболочки от рептилий. У яйцекладущих однопроходных эти оболочки все еще выполняют свои предковые функции, поскольку энергетические потребности эмбриона удовлетворяются богатыми запасами желтка в крупных яйцах, покрытых скорлупой. У зародышей сумчатых и плацентарных, которые получают большую часть необходимой для развития энергии от матери, яйца содержат мало желтка, и эмбрион вскоре прикрепляется к стенке матки с помощью проникающих в нее выростов хориона. У большинства сумчатых и у некоторых плацентарных он сливается с желточным мешком и формирует примитивную плаценту, называемую желточной. Плацента (называемая также детским местом или последом) – это образование, которое обеспечивает двусторонний обмен веществ между эмбрионом и материнским организмом. Через нее происходит поступление питательных веществ к зародышу, его дыхание и удаление продуктов обмена. У большинства плацентарных млекопитающих хорион образует ее вместе с аллантоисом, и она называется аллантоидной.
Продолжительность периода от оплодотворения яйцеклетки до рождения детеныша варьирует от 12 дней у некоторых сумчатых до примерно 22 месяцев у африканского слона. Число новорожденных в помете обычно не превышает числа сосков у матери и, как правило, меньше 14. Обычно из оплодотворенной яйцеклетки развивается один зародыш, но встречается и полиэмбриония, т.е. она дает несколько эмбрионов, разделяющихся на самых ранних стадиях развития. Изредка это происходит у многих видов, в том числе у человека рождаются полностью идентичные – однояйцевые индивиды – близнецы.
Непосредственно после рождения (у однопроходных, после вылупления из яиц) млекопитающие питаются материнским молоком. Молочные железы обычно расположены парами. В то же время у многих сумчатых число молочных желез нечетное и развит только один сосок посередине живота.
В целом механизм передвижения (локомоции) одинаков у всех млекопитающих, но конкретные его способы развивались по многим расходящимся направлениям.
Когда предки зверей впервые выползли на сушу, их передние и задние конечности были короткими и широко расставленными, что делало передвижение по земле медленным и неуклюжим. Эволюция способов локомоции млекопитающих была направлена в основном на увеличение скорости путем удлинения и выпрямления ног и поднятия туловища над землей. Этот процесс требовал определенных изменений скелета, включая утрату ряда элементов рептильного плечевого пояса. Благодаря разнообразию специализации звери освоили все возможные экологические ниши. У современных млекопитающих способы локомоции включают рытье, ходьбу, бег, прыжки, лазание, планирование, машущий полет и плавание. Роющие формы, например кроты, движутся под поверхностью почвы. Мощные передние конечности этих млекопитающих выдвинуты вперед, так что лапы могут работать перед головой, а плечевые мышцы очень сильно развиты. В то же время их задние конечности слабые и неспециализированные. Кисти таких зверей могут быть очень большими, приспособленными для разгребания мягкой почвы или же вооруженными мощными когтями для «бурения» твердого грунта.
Многие мелкие виды, например крысы, мыши и землеройки, характеризуются сравнительно массивным туловищем с короткими конечностями и обычно передвигаются перебежками. Говорить о какой-то их локомоторной специализации вряд ли стоит. Некоторые млекопитающие, например медведи, лучше всего приспособлены к ходьбе. Они принадлежат к стопоходящему типу и опираются при ходьбе на стопы и ладони. В случае необходимости они могут переходить на тяжелый бег, но делают это неуклюже и не могут долго сохранять высокую скорость. Приспособлены к ходьбе также очень крупные животные, например слоны, у которых наблюдается тенденция к удлинению и усилению верхних костей ног при укорочении и расширении нижних. Это превращает конечности в массивные колонны, поддерживающие огромную массу тела. И, напротив, у быстро бегающих животных типа лошадей и оленей нижние сегменты ног стержневидные, способные быстро двигаться вперед и назад. Мышцы конечностей при этом сконцентрированы в их верхней части, оставляя внизу в основном мощные сухожилия, скользящие, как по блокам, по гладким поверхностям хряща и тянущиеся до мест прикрепления к костям стоп и кистей. Дополнительные приспособления к быстрому бегу включают редукцию или утрату наружных пальцев и сближение оставшихся.
Необходимость догонять проворную добычу и в кратчайшие сроки преодолевать большие расстояния, ведя ее поиск, привела к появлению у кошек и собак другого способа локомоции – на пальцах. Пясть и плюсна при этом удлинились, что позволило увеличить скорость бега. Ее рекорд для млекопитающих зафиксирован у гепардов: примерно 112 км/ч.
Другим основным направлением эволюции быстрого передвижения по земле стало развитие способности к прыжкам. Большинство животных, жизнь которых находится в прямой зависимости от скорости их локомоции, движется вперед, используя в основном толчки задних ног. Крайнее развитие этого способа передвижения, сочетавшееся со сменой образа жизни, привело к глубоким структурным преобразованиям прыгающих видов. Главным их морфологическим изменением было удлинение задних конечностей, в первую очередь их нижних отделов, приведшее к усилению толчка и способности смягчать удар при приземлении. Для обеспечения силы, необходимой для длинных следующих друг за другом скачков, мышцы этих конечностей сильно разрослись в поперечном направлении. Одновременно их наружные пальцы редуцировались или вообще исчезли. Сами конечности широко раздвинулись для увеличения устойчивости, и животное в целом стало пальцеходящим. В большинстве случаев передние конечности сильно уменьшились, а шея укоротилась. Хвост таких видов очень длинный, как у тушканчика, или сравнительно короткий и толстый, как у кенгуру. Он служит балансиром и до некоторой степени рулевым устройством.
Прыжковый способ локомоции позволяет добиться максимального ускорения. Вычисления показывают, что самый длинный прыжок возможен при угле отрыва от земли, равном 40–44○. Кролики используют способ передвижения, промежуточный между бегом и прыжками: мощные задние ноги толкают тело вперед, но приземляется животное на передние лапы и готово повторить прыжок, только вновь сгруппировавшись в исходное положение.
Чтобы удлинить прыжки и, таким образом, более эффективно преодолевать расстояние, некоторые звери приобрели перепонку, тянущуюся вдоль тела между передними и задними конечностями и прикрепленную к запястьям и лодыжкам. При раскрывании конечностей она расправляется и обеспечивает достаточную подъемную силу для планирования сверху вниз между расположенными на разной высоте ветвями. Грызун летяга – типичный пример животных, передвигающихся таким способом. Сходные перепонки для планирования независимо развились в других группах, в том числе у африканских шипохвостов и австралийской сумчатой летяги (летающего опоссума). Начать полет животное может практически из любого положения. Вытянув голову вперед, оно скользит по воздуху, набирая под действием силы тяжести скорость, достаточную для разворачивания тела вверх перед посадкой, так что идет на нее в вертикальном положении. После этого животное готово карабкаться по стволу дерева вверх и, забравшись на необходимую высоту, повторить полет.
За исключением такого планирования, которое эволюционировало как особый тип локомоции, у современных млекопитающих не наблюдается никаких переходов от передвижения по земле к машущему полету.
Единственные млекопитающие, на самом деле способные летать, – рукокрылые. Древнейшие из известных ископаемых представителей уже имели хорошо развитые крылья, строение которых за 60 млн. лет почти не изменилось. Предполагается, что эти летающие млекопитающие произошли от какой-то примитивной группы насекомоядных.
Характер полета рукокрылых разных родов и видов неодинаков. Некоторые из них, например, крылановые, размеренно взмахивают крыльями. Складчатогубы летают очень быстро, а скорость полета может резко меняться. Некоторые летают плавно, как ночные бабочки. Как бы там ни было, полет – главный способ локомоции рукокрылых, некоторые мигрирующие виды покрывают без отдыха до нескольких сотен километров.
Хотя бы один представитель почти каждого отряда млекопитающих хорошо плавает. Фактически, все звери, даже рукокрылые, могут в случае необходимости держаться на воде. Ленивцы передвигаются в ней даже быстрее, чем на суше, а некоторые кролики освоили эту среду не хуже ондатр. Существуют различные уровни специальной адаптации млекопитающих к жизни в воде. Например, у норки нет для этого никаких особых приспособлений, за исключением смазанного жиром меха, а киты формой тела и поведением напоминают скорее рыб, чем зверей. У полуводных форм задние лапы обычно увеличены и снабжены перепонкой между пальцами или оторочкой из жестких волос, как у выдры. Их хвост может быть преобразован в гребную лопасть или руль, став уплощенным вертикально, как у ондатры, или горизонтально, как у бобра. В то же время они все еще сохраняют толстый мех для сохранения тепла и способны передвигаться по суше на четырех конечностях.
Полное приспособление к жизни в воде демонстрируют китообразные и сирены. Оно сопровождается глубокими морфологическими преобразованиями, включая полное исчезновение наружных задних конечностей, приобретение обтекаемой, рыбообразной формы тела и исчезновение волосяного покрова. Сохранять тепло китам, как и настоящим тюленям, помогает толстый слой ворвани, окружающий тело. Поступательное движение в воде обеспечивают горизонтальные плавники с хрящевым каркасом, расположенные в задней части хвоста.
Все млекопитающие развили определенные механизмы самосохранения, причем многие в ходе эволюции приобрели особые защитные приспособления.
Защитные покровы. Некоторые звери, например еж, покрыты иглами и в случае опасности сворачиваются клубком, выставляя их во все стороны. Сходный способ защиты применяют броненосцы, способные полностью отгораживаться от окружающего мира роговым панцирем, который предохраняет тело и от острых колючек кактусов, представляющих собой наиболее распространенную растительность в местах обитания этих животных. Североамериканский дикобраз пошел в развитии защитных покровов еще дальше. Он не просто покрыт зазубренными иглами, которые, застряв в теле врага, могут привести к его гибели, но и очень ловко орудует колючим хвостом, нанося им быстрые и точные удары по противнику.
Железы. Млекопитающие используют для защиты и химическое оружие. Этот метод наиболее освоен скунсом, который вырабатывает в парных анальных железах у основания хвоста едкую и очень вонючую жидкость. Сокращая окружающие железы мышцы, он может выбрасывать ее тонкую струю на расстояние до 3 м, целясь в наиболее уязвимые места противника – глаза, нос и рот.
Кератин является важной составляющей наружного слоя кожи (эпидермиса) млекопитающих. Это прочный, эластичный и нерастворимый в воде белок. Он крайне необходим для защиты животных, поскольку предохраняет нижележащие ткани от химических раздражителей, влаги и механических повреждений. Участки кожи, особо подверженные агрессивному действию внешней среды, защищены утолщенным эпидермисом с увеличенным содержанием кератина. Примером могут служить мозолистые наросты на подошвах. Когти, ногти, копыта и рога – все это специализированные кератиновые образования.
Когти, ногти и копыта состоят из одних и тех же структурных элементов, но различаются по их расположению и степени развития. Коготь состоит из двух частей – верхней пластинки, называемой когтевой, и нижней подошвенной. У рептилий они обычно образуют две половины конического колпачка, охватывающего мясистый конец пальца. В когтях млекопитающих нижняя пластинка уменьшена и палец практически не покрывает. Верхняя пластинка ногтя широкая и плоская, а узкий остаток нижней скрыт между ее краем и подушечкой пальца. В копыте обе пластинки увеличены, утолщены и изогнуты, причем верхняя (стенка копыта) окружает нижнюю (его подошву). Мясистый конец пальца, называемый у лошадей стрелкой, оказывается при этом оттесненным назад и вверх.
Когти используются в основном для рытья, лазания и нападения. Бобр раздвоенным когтем задней лапы расчесывает мех. Кошки обычно держат когти втянутыми в специальные чехлы, чтобы не затупить их концы. Олени часто защищаются острыми, как топор, копытами и могут убивать ими змей. Лошадь славится мощным ударом задних ног, причем способна лягаться каждой ногой в отдельности и обеими сразу. Защищаясь, она может также встать на дыбы и резко ударить противника сверху вниз передними копытами.
Рога. В процессе эволюции млекопитающие очень рано приобрели выросты черепа, используемые в качестве оружия. Они были у некоторых видов уже в эоцене (ок. 50 млн. лет назад) и становились с тех пор все более характерными для многих копытных. В плейстоцене (начался ок. 1,6 млн. лет назад) эти выросты достигали фантастических размеров. Во многих случаях они важнее для драк с сородичами, например при конкуренции самцов за самку, чем как средство защиты от хищников.
В принципе все рога – это твердые выросты на голове. Однако развивались и специализировались они в двух разных направлениях. Один тип можно назвать настоящими рогами. Они состоят из отходящей от лобных костей обычно не разветвленной костной сердцевины, покрытой чехлом из твердой кератинизированной роговой ткани. Этот снятый с черепных выростов полый чехол используется для изготовления различных «рогов», в которые трубят, наливают вино и т.п. Настоящие рога обычно присутствуют у животных обоих полов и не сбрасываются в течение всей их жизни. Исключение составляют рога американского вилорога. Их роговой, как у настоящих рогов, чехол не только несет небольшой отросток (иногда не один), образуя «вилку», но и каждый год сбрасывается (сменяется).
Второй тип – это рога оленей, которые в полностью развитом виде состоят только из кости без рогового покрытия, т.е. собственно «рогами» их называют неправильно. Это тоже отростки лобных костей черепа, обычно ветвистые. Рога оленьего типа присутствуют только у самцов, хотя и здесь исключение составляет северный олень. В отличие от настоящих, такие рога каждый год сбрасываются и отрастают заново.
Настоящие рога характерны для группы полорогих – крупного рогатого скота, овец, коз и антилоп. У диких буйволоподобных млекопитающих они зачастую сильно утолщены у основания и образуют как бы шлем, например у овцебыка и черного африканского буйвола. У большинства типов крупного рогатого скота они лишь слегка изогнуты. Концы рогов у всех видов в той или иной степени направлены вверх, что увеличивает их эффективность в качестве оружия.
Рога снежного барана по отношению к общим размерам животного самые тяжелые и крупные. У самцов они массивные и закручены в спираль, меняющую свою форму в ходе роста, так что их концы могут в итоге описать более одного полного круга. В бою эти рога используются как таран, а не в качестве колющего оружия. У самок они мельче и почти прямые.
Рога диких козлов специализировались иначе. Внушительность им придает длина. Дуговидные, широко расходящиеся у горного козла и прямые, закрученные штопором у козла винторогого, они сильно отличаются от бараньих, которые даже при большей общей длине кажутся мельче, поскольку их концы из-за спирального изгиба находятся ближе к основанию.
Зубы. У большинства безрогих млекопитающих основное оружие – зубы. Впрочем, некоторые виды, например муравьеды, их лишены, а, скажем, кролики с прекрасно развитыми зубами никогда не используют их для защиты, как бы велика ни была опасность. Большинство грызунов в случае угрозы находит достойное применение своим долотовидным резцам. Летучие мыши могут кусаться, но в большинстве случаев их зубы слишком малы для нанесения серьезных ран. Хищные используют в бою главным образом острые, длинные клыки, которые им жизненно необходимы. Кошачьи клыки опасны, но укус собак мощнее, поскольку в поединке эти животные не способны помогать себе когтями.
Некоторые млекопитающие приобрели в ходе эволюции высокоспециализированные зубы, называемые бивнями. Они используются в основном для добывания пищи, но могут служить и оружием. Большинство диких свиней, например европейский дикий кабан, своими длинными клыками-бивнями выкапывают съедобные коренья, но могут с помощью этих зубов нанести и серьезную рану противнику.
Африканские слоны – обладатели самых крупных бивней среди ныне живущих млекопитающих. Они используют их в бою, для рытья и мечения территории. Пара таких бивней может достигать общей длины 3 м, давая более 140 кг слоновой кости.
По агрессивности поведения млекопитающих можно разделить на три основные группы: безобидные (никогда не нападающие на теплокровных животных с целью убийства), безразличные (способные на спровоцированные нападение и убийство) и агрессивные (убивающие регулярно).
Безобидные. Кролики, возможно, наиболее безобидные из всех млекопитающих: они не пытаются даже сделать вид, что дерутся, каким бы отчаянным ни было их положение. Грызуны в целом безобидны, хотя некоторые виды, например обыкновенная белка, может при случае убить и съесть мелкого зверька. Синий кит – самое крупное и сильное из когда-либо живших млекопитающих, однако питается он мелкими ракообразными и рыбешками, относясь, таким образом, к безобиднейшим существам.
Дата добавления: 2015-08-21; просмотров: 53 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Лабораторна робота №1 | | | Выезд из Львова 8 дней |