Глюконеогенез – це процес синтезу глюкози з невуглеводних субстратів. Такими попередниками глюкози є лактат, піруват, більшість амінокислот, гліцерин, проміжні продукти циклу лимонної кислоти. Відбувається глюконеогенез у печінці й, невеликою мірою, в кірковій речовині нирок. Завдяки цьому процесу підтримується концентрація глюкози в крові після того, як вичерпаються запаси глікогену при вуглеводному чи повному голодуванні. Надзвичайно важливе значення глюконеогенезу для організму тварин і людини зумовлюється тим, що мозок має дуже малі запаси глікогену і глюкоза крові служить основним джерелом енергії для нього. При зменшенні концентрації глюкози в крові нижче певної критичної межі порушується функціонування мозку і може настати смерть. Механізм глюконеогенезу також забезпечує видалення з крові таких продуктів тканинного метаболізму, як лактат і гліцерин.
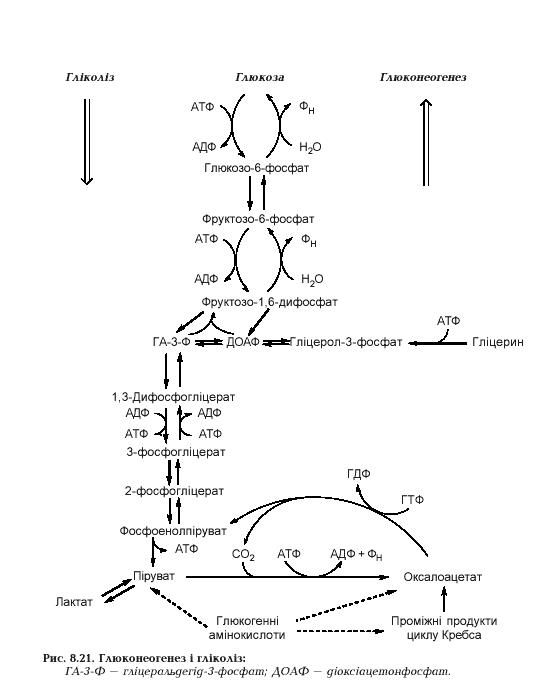
Процес зворотний до гліколізу, але три реакції гліколізу є фактично незворотними, тому глюконеогенез включає 7 гліколітичних реакцій, які мають зворотний напрямок, і три обхідні стадії (рис.).
http://www.youtube.com/watch?v=p-lFJVOkFwk
Обхідні реакції каталізуються іншими ферментами і є також незворотними, але вони йдуть у напрямку синтезу глюкози.
Перша незворотна стадія – перетворення пірувату у фосфоенолпіруват, сполуку з макроергічним зв'язком. Здійснюється воно через проміжну речовину – оксалоацетат. Спочатку мітохондріальний фермент піруваткарбоксилаза каталізує АТФ-залежне карбоксилювання пірувату:

Алостеричним активатором піруваткарбоксилази служить ацетил-КоА. Як розглянуто раніше, піруваткарбоксилазна реакція також постачає оксалоацетат для циклу лимонної кислоти.
У глюконеогенезі оксалоацетат під дією фосфоенолпіруват-карбоксикінази перетворюється у фосфоенолпіруват:
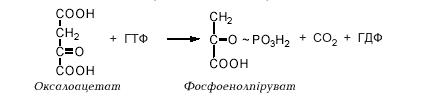
Одночасно відбуваються декарбоксилювання і фосфорилювання. ГТФ, який використовується в цій реакції, регенерується шляхом взаємодії ГДФ з АТФ. Таким чином, на перетворення пірувату у фосфоенолпіруват витрачаються 2 молекули АТФ, тоді як у процесі гліколізу при перетворенні фосфоенолпірувату в піруват синтезується тільки 1 молекула АТФ. Труднощі викликає різна локалізація ферментів у клітині. Фосфоенолпіруват-карбоксикіназа знаходиться в цитоплазмі, а оксалоацетат, який утворюється в мітохондріях, не проникає через мітохондріальну мембрану. Тому оксалоацетат відновлюється в матриксі мітохондрій під дією ферменту циклу Кребса малатдегідрогенази до малату, який може виходити з мітохондрій. Цитоплазматична малатдегідрогеназа окиснює малат назад в оксалоацетат.
Утворений фосфоенолпіруват далі переходить за допомогою зворотних реакцій гліколізу у фруктозо-1,6-дифосфат. На етап перетворення 3-фосфогліцерату в 1,3-дифосфогліцерат затрачається одна молекула АТФ. Фосфофруктокіназна реакція гліколізу фруктозо-6-фосфат – фруктозо-1,6-дифосфат незворотна, тому інший фермент (фруктозо-дифосфатаза) каталізує гідроліз фруктозо-1,6-дифосфату до фруктозо-6-фосфату:

Вивільняється неорганічний фосфат, а енергія макроергічного фосфодіефірного зв'язку виділяється у вигляді тепла. У наступній зворотній реакції фруктозо-6-фосфат перетворюється в глюкозо-6-фосфат, який в обхідній реакції під дією глюкозо-6-фосфатази гідролізується до вільної глюкози:
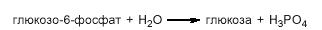
Крім того, глюкозо-6-фосфат може використовуватись для синтезу глікогену.
Враховуючи необхідність двох молекул пірувату для утворення молекули глюкози, сумарне рівняння глюконеогенезу записується так:
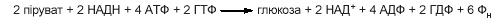
Таким чином, на синтез глюкози з пірувату затрачається значно більше енергії, ніж її утворюється під час гліколізу (шість молекул АТФ проти двох).
Для синтезу глюкози шляхом глюконеогенезу використовуються проміжні продукти циклу лимонної кислоти, які перетворюються в циклі в оксалоацетат. Фактичними субстратами є амінокислоти, які після втрати аміногруп стають проміжними продуктами циклу Кребса та піруватом. Такі амінокислоти називаються глюкогенними. Глюконеогенез з амінокислот інтенсивно відбувається при голодуванні та цукровому діабеті. Крім того, в цих умовах розпадаються жири жирової тканини, причому жирні кислоти застосовуються як джерело енергії в м'язах, печінці й інших тканинах, а гліцерин у печінці шляхом глюконеогенезу переходить у глюкозу.
Іншим важливим субстратом глюконеогенезу є молочна кислота, яка накопичується в організмі під час інтенсивної м'язової роботи внаслідок анаеробного розпаду глікогену. У період відновлення після напруженої роботи молочна кислота переноситься кров'ю з м'язів до печінки, де під дією лактатдегідрогенази окиснюється до пірувату. Частина останнього використовується для глюконеогенезу, а частина розпадається аеробним шляхом, забезпечуючи процес глюконеогенезу АТФ. Глюкоза потрапляє назад у скелетні м'язи і застосовується для відновлення запасу глікогену. Поєднання процесу анаеробного гліколізу в скелетних м'язах і глюконеогенезу в печінці називається циклом Корі й зображене на рис.
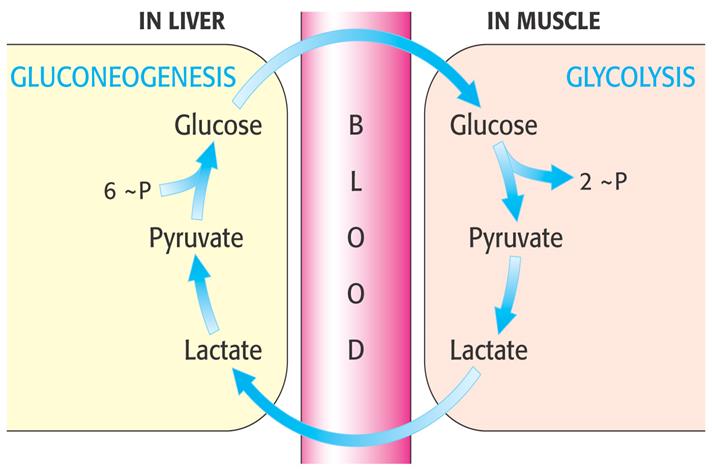

Молочна кислота утворюється постійно в еритроцитах, мозковій частині нирок, сітківці ока, а в печінці й корі нирок переходить у глюкозу, яка повинна знову надходити в названі клітини і використовуватись. Таким чином, ця кислота, на відміну від глюкогенних амінокислот, не служить попередником для глюкози крові, що могла б використовуватись у мозку і нервах при голодуванні.
http://www.youtube.com/watch?v=qF3ylhC0VeQ
Регуляція глюконеогенезу
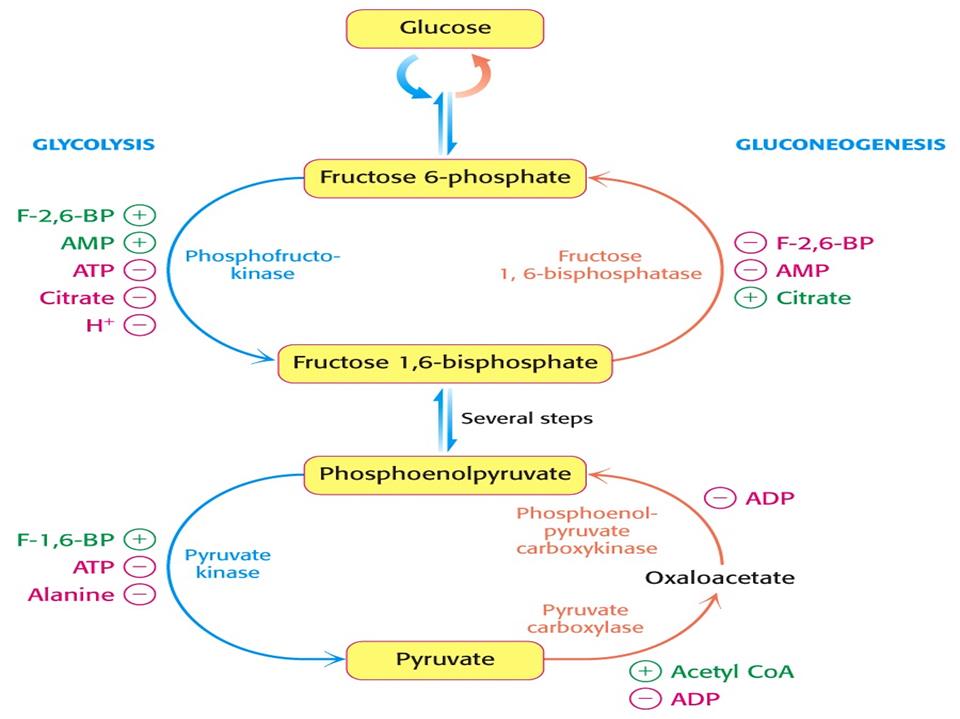
У клітинах печінки здійснюється координована регуляція гліколізу і глюконеогенезу відповідно до фізіологічних потреб усього організму. Система контролю включає субстрати і проміжні продукти процесів, регуляторні ферменти та їх ефектори, гормони. Підкреслимо, що регуляторними ферментами глюконеогенезу і гліколізу є ті, що каталізують незворотні реакції і не беруть участі у протилежному процесі. Наприклад, піруваткарбоксилаза (активатор – ацетил-КоА) і фруктозодифосфатаза (активатор – цитрат, інгібітори – АМФ і фруктозо-2,6-дифосфат). На рис. показані регуляторні пункти гліколізу і глюконеогенезу та контрольні чинники.

Деякі ефектори одночасно впливають на активність регуляторних ферментів в обох процесах. Так, ацетил-КоА служить активатором піруваткарбоксилази і інгібітором піруваткінази. Крім того, ацетил-КоА є інгібітором піруватдегідрогеназного комплексу і, сповільнюючи розпад пірувату до ацетил-КоА, сприяє переходу пірувату в глюкозу. АМФ інгібує фруктозо-дифосфатазу і разом з тим активує відповідний фермент гліколізу – фосфофруктокіназу. Навпаки, цитрат – активатор фосфатази й інгібітор кінази.
Глюконеогенез стимулюється не тільки активаторами піруваткарбоксилази і фруктозодифосфатази, а й інгібіторами регуляторних ферментів гліколізу (АТФ, аланін, жирні кислоти). Таким чином, коли в клітині є достатня концентрація палива для циклу лимонної кислоти (ацетил-КоА, оксалоацетату, цитрату, жирних кислот, аланіну) чи висока концентрація АТФ і низькі –АДФ та АМФ, посилюється біосинтез глюкози і гальмується гліколіз. У протилежній ситуації стимулюється гліколіз і гальмується глюконеогенез.
На глюконеогенез впливають такі гормони, як глюкагон й інсулін підшлункової залози та глюкокортикоїди кори надниркових залоз. При голодуванні глюкагон посилює розпад жирів у жировій тканині. Жирні кислоти надходять у печінку, де розпадаються до ацетил-КоА. Швидкість окиснення ацетил-КоА в циклі лимонної кислоти відстає від швидкості його утворення, і підвищений рівень ацетил-КоА в клітині активує піруваткарбоксилазу. В результаті посилюється глюконеогенез.
http://www.youtube.com/watch?v=6fmYLF-uiNI
Глюкагон започатковує глюконеогенез ще одним способом – через фруктозо-2,6-дифосфат. Цей регуляторний компонент відкритий у 1980 р. Утворюється фруктозо-2,6-дифосфат із фруктозо-6-фосфату при дії фосфофруктокінази ІІ, а розщеплюється фруктозо-2,6-дифосфатазою:
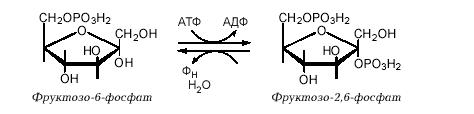
Фруктозо-2,6-дифосфат активує фермент гліколізу фосфофруктокіназу і гальмує активність ферменту глюконеогенезу фруктозо-1,6-дифосфатази. Утворення фруктозо-2,6-дифосфату пригнічується глюкагоном, який шляхом цАМФ-залежного фосфорилювання інактивує фосфофруктокіназу ІІ й активує фруктозо-2,6-дифосфатазу. Таким чином, при голодуванні глюкагон знижує внутрішньоклітинну концентрацію фруктозо-2,6-дифосфату, що викликає стимуляцію глюконеогенезу і гальмування гліколізу. Підвищення швидкості глюконеогенезу в печінці призводить до того, що глюкоза надходить у кров і потрапляє в інші органи, насамперед у мозок.
Субстратами глюконеогенезу служать амінокислоти, оскільки під час голодування в крові низьке співвідношення інсулін/глюкагон гальмує синтез білків і стимулює їх катаболізм, зокрема в м'язах, що забезпечує постачання амінокислот у печінку. При тривалому голодуванні зростає секреція корою надниркових залоз глюкокортикоїдів, які посилюють у печінці синтез ферментів глюконеогенезу (фосфоенолпіруват-карбоксикінази, глюкозо-6-фосфатази) й амінотрансфераз – ферментів, які каталізують перетворення глюкогенних амінокислот у піруват і оксалоацетат. У м'язах та інших тканинах глюкокортикоїди гальмують синтез білків. У результаті стимуляції глюконеогенезу глюкокортикоїди збільшують концентрацію глюкози в крові й синтез глікогену в печінці. Інсулін протидіє стимулюючій дії глюкагону і глюкокортикоїдів на глюконеогенез.
Дата добавления: 2015-09-05; просмотров: 330 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Обмін глікогену | | | РЕГУЛЯЦІЯ РІВНЯ Глюкози в крові |