Читайте также:
|
Как указывалось в §5.1, биологические системы, в том числе, естественно, клетки представляют собой двухступенчатую каскадную управляемую систему, у которой первая ступень обеспечивает устойчивое сохранение информации в эталонном органе – ДНК, а вторая – устойчивое сохранение порядка во всей системе в целом (в данном случае – сохранение циклических химических реакций полимеризации белков и нуклеиновых кислот в клетке). В связи с этим удобно разделить рассмотрение этих процессов также на две части – сначала изучить процессы сохранения наследственной информации (будем именовать их далее «информационными процессами»), а затем процессы сохранения информации, передаваемой от ДНК другим органам клетки – процессы жизнедеятельности клетки. Эту вторую часть обычно именуют процессами управления (регуляции) клетки и многоклеточного организма.
В свою очередь процессы управления жизнедеятельностью биосистем будут рассматриваться отдельно на уровне клеток и на уровне многоклеточных живых организмов в следующих параграфах.
Информационные процессы в клетке подразделяются на следующие виды:
1)процессы формирования клеточных структур на базе наследственной информации;
2)процессы, обеспечивающие сохранение наследственной информации;
3)процессы передачи наследственной информации при делении клетки (митозе);
4)процессы передачи наследственной информации в многоклеточных организмах;
5)процессы изменения наследственной информации в многоклеточных организмах.
Рассмотрим все эти процессы последовательно. В цепочках молекул ДНК закодированы все типы молекул белка данного организма. Программа кодирования следующая. Каждым трем расположенным рядом узлам одной из нитей спирали ДНК соответствует одна молекула аминокислоты (из 20 применяемых в биосистемах). В таблице 5.1 показано, какой тройке нуклеотидов соответствует какая аминокислота. Как видим, тройке нуклеотидов АГГ соответствует аминокислота арг, тройке ААГ – лиз и т.д. Из таблицы видно, что одна и та же аминокислота может быть закодирована несколькими тройками нуклеотидов. Так, аминокислота про соответствует тройкам ЦЦА, ЦЦГ, ЦЦЦ, и ЦЦТ, а цис – ТГЦ и ТГТ. Тройки нуклеотидов называются кодонами.
Таблица 5.1
| А | Г | ||||||||||||
| А | Г | Т | Ц | А | Г | Т | Ц | ||||||
| АГ | ТЦ | АГ | ТЦ | Г | АТЦ | АГТЦ | АГ | ТЦ | АГТЦ | АГТЦ | АГТЦ | ||
| лиз | асн | арг | сер | мет | иле | тре | глу | асп | гли | вал | ала | ||
| Т | Ц | ||||||||||
| А | Г | Т | Ц | А | Г | Т | Ц | ||||
| АГ | ТЦ | А | Г | ТЦ | АГТЦ | АГТЦ | АГ | ТЦ | АГТЦ | АГТЦ | АГТЦ |
| тер | тир | тер | три | цис | лей,фен | сер | гл | гис | арг | лей | про |
Символ тер в таблице 5.1 означает терминирующий кодон, т.е. такую тройку нуклеотидов, которая соответствует концу строящейся молекулы РНК. Начало строительства РНК определяется кодоном АТГ и ГТГ, если перед этим была пауза. Эти кодоны именуются иницирующими.
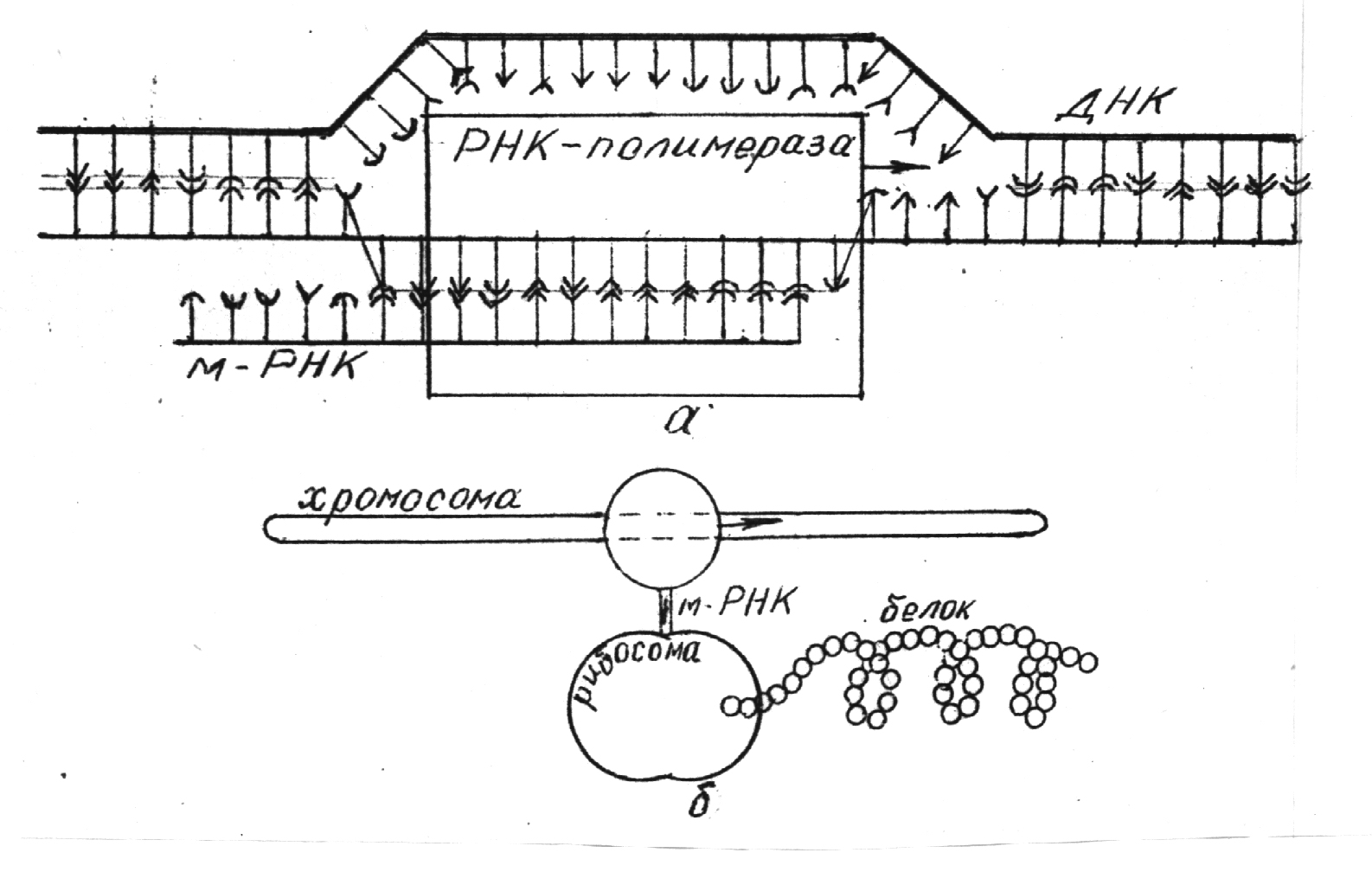
Процессу производства белка у простейших бактерий соответствует рисунок 5.7. Вдоль молекулы ДНК внутри хромосомы скользит фермент РНК-полимераза, расщепляя двойную цепь на две одномерные (рисунок 5.7,а). На одной из этих двух нитей ДНК в соответствии с правилом комплементарности (нуклеотиду А нити ДНК соответствует нуклеотид У РНК, Г соответствует Ц, Т – А, а Ц – Г) формируется нить мРНК. В качестве строительного материала используются нуклеотиды РНК-полимеразы. Далее одномерная цепочка мРНК входит внутрь рибосомы (рисунок 5.7,б), где на каждом кодоне формируется узел белка – аминокислотный остаток. Аминокислотный остаток доставляется в рибосому транспортной РНК (тРНК), которая имеет два свободных конца с тремя соответствующими нуклеотидами: если на одном конце тройка УГЦ, то на другом АЦГ, т.е. парная первой. К первому концу прикрепляется соответствующий пептид (в данном случае цис), а другой присоединяется к соответствующему участку мРНК (т.е. к участку с кодоном УГЦ). Пептид “приклеивается” – присоединяется к формирующейся цепочке белка, после чего данная молекула тРНК отделяется от мРНК и выходит из рибосомы. Процесс этот идет довольно быстро – 20 узлов в секунду. Как видим, мРНК считывает информацию, закодированную в ДНК и синтезирует соответствующий белок. Подходя к кодону УАГ, цепочка матричной РНК прерывается. Соответствующая молекула белка готова. Участок ДНК, с которой копируется данная молекула белка, именуется геном.
Рисунок 5.7.
Схема строительства матричной(информационной)РНК (а)и изготовления белка в рибосоме(б).
У высокоорганизованных животных и растений цепочка РНК, кодирующая ген, снимается с очень длинного участка ДНК и включает в себя посторонние нуклеотиды, не имеющие отношения к данному гену.Ген оказывается в цепочке ДНК расчленен на части, между которыми расположены участки посторонних кодонов, не имеющих отношения к данному белку. По сути дела, ситуацию можно проиллюстрировать на примере такой записи файла на дискете компьютера, при которой его программа разорвана на части, между которыми включены участки других файлов. Вместо того, чтобы сразу "штамповать" белок, полученная молекула РНК (именуемая про-РНК) разрезается специальным белком-ферментом – рестриктазой – на участки, из которых нужные (экзоны) снова сшиваются, а ненужные (интроны) – выбрасываются. Процесс этот именуется сплайсингом.
Следующий процесс, связанный с ДНК и передачей наследственных признаков – это процесс удвоения ДНК при делении клеток – репликация. Молекулы ДНК в этом процессе служат основой для передачи наследственной информации.
Осуществляется репликация следующим образом. На определенной стадии эволюции клетки, незадолго до процесса деления под воздействием находящихся в хромосоме белков-ферментов молекула ДНК расщепляется на две нити – рисунок 5.8. К каждой нити, как к матрице, присоединяются соответствующие нуклеотиды, которые, соединяясь между собой, образуют две новые нити. Так формируется пара новых молекул ДНК, являющихся точной копией исходной. Возможность деления молекулы ДНК на две – одна из основных функций, обеспечиваемых наличием двух нитей (комплементарностью ДНК).
Вторая функция – это ремонт повреждений молекулы ДНК, происходящих под воздействием внешних факторов – ультрафиолетового и радиоактивного облучений, излучения Вавилова-Черенкова, случайно попадающих в цитоплазму посторонних химических веществ и микробиологических объектов – репарация.
Рисунок 5.8 Процесс удвоения молекулы ДНК
В многоклеточных, особенно в сложных высокоразвитых организмах, процесс сохранения наследственной информации обеспечивается также механизмом полового размножения.
В этих организмах передача информации от одного поколения биосистем следующему (дочернему) осуществляется специальными (половыми) клетками организма. Все остальные клетки (соматические) заняты обеспечением жизнедеятельности организма. При этом они делятся, увеличивая размеры того или иного органа или количество жизненно важного продукта (кровь, гормоны и т.п.), погибают (прекращают деление) или вообще не делятся, сохраняясь неизменными в течение всей жизни организма.
Половые клетки способны начать воспроизводство дочерних особей лишь после слияния (сингамии) двух клеток, происходящих от разных родителей. При этом различаются как сами родители (мужская и женская особи), так и строение и функция самих клеток. Такой механизм возник в природе для обеспечения целенаправленного и достаточно гибкого изменения наследственной информации. Благодаря этому процессу возникает дополнительный контроль точности сохранения основных элементов наследственной информации, так как дочерний организм появляется лишь от клеток особей, принадлежащих к одному и тому же виду.
Изменения организма касаются лишь второстепенных признаков и приводят к качественному изменению данного вида лишь в результате постепенного накопления от поколения к поколению. Ясно, что благодаря диплоидному размножению одновременно происходит "опрос" максимального числа особей. Пусть, например, качественное изменение вида произошло через 100 поколений. Ясно, что при этом произошел опрос 2100=1030 особей. Конечно, родительские особи могут состоять в родстве, т.е. иметь общих предков. В этом случае количество "опрошенных" сокращается. Зато происходит усиленное развитие признака, присущего “родственникам”. Поскольку, как правило, диплоидное размножение происходит между особями, территориально связанными друг с другом, на возникновение и усиление указанного признака оказывают преимущественное влияние условия среды. Так внутри популяции (вида) возникает подвид, а из него в процессе эволюции – новый вид. Процесс накопления определённого признака путём усиления влияния какого-либо гена из уже существующих в данном виде достаточно хорошо изучен и будет описан ниже. Гораздо сложнее выявить процесс нарождения новых генов. Однако совершенно ясно, что механизм их случайного появления вследствие мутаций молекулы ДНК, вызванных неблагоприятным воздействием окружающей среды, не может привести к созданию более совершенных и структурно более сложных живых организмов. В последние годы появились новые данные (в частности, при изучении механизма иммунной системы животных), свидетельствующих о наличии целенаправленного механизма “конструирования” новых генов [20].
Рассмотрим, как возникающие при жизни одного организма изменения передаются от поколения к поколению.
В диплоидном организме за формирование того или иного органа – носа, волос, позвонков и т.д. – отвечают два гена – два участка ДНК, расположенных в гомологичных (полученных от мужской и женской половой клетки) хромосомах (аллели). При этом каждый из них отличается друг от друга второстепенными признаками: например, цветом волос, формой носа и т.д. Именно это обеспечивает своеобразие каждого организма – ни один человек не похож на другого, ни один цветок не повторяет в точности другой.
При образовании дочернего организма он усваивает один из признаков родителей. При этом оказывается, что признаки обоих родителей не равноценны - один из них явно доминирует над другим. Этот признак называется доминантным в отличие от второго – рецессивного. В том случае, если гены обоих родителей обладают доминантным признаком (например, карими глазами), то и у потомка будет этот же признак – тоже карие глаза. Если у одного из родителей доминантный признак, а у другого рецессивный, то у потомка вновь будет доминантный, хотя он содержит смесь двух генов. Если один родитель содержит смесь доминантного и рецессивного гена, а другой – только рецессивный, то у потомка может быть либо набор только рецессивных генов, либо смешанный набор. Если у обоих родителей смешанные наборы, то у потомка возможны три варианта – чисто доминантный набор, чисто рецессивный набор и смешанный.
Количество признаков одного и того же органа может быть не два, а больше – три, пять, шесть... В этом случае они будут находиться друг по отношению к другу в сложной зависимости доминантности и рецессивности. При этом могут возникнуть промежуточные признаки – пятнистая раскраска шерсти, промежуточная окраска цветка и т.п. На доминантность того или иного аллеля могут влиять другие гены, зашифрованные в соответствующей молекуле ДНК. Так обеспечивается разнообразие признаков и различие особей внутри одного вида.
Еще более усложняется процесс размножения признаков в результате кроссинговера – частичной передачи части гомологичных молекул ДНК в период созревания (мейоза) половых клеток. Вследствие этого взаимные влияния генов друг на друга еще более запутываются. Одновременно ряд признаков из доминантных переходят в рецессивные.
До сих пор речь шла о признаках, так или иначе унаследованных от далеких предков. Все указанные комбинации лишь увеличивают число вариантов признаков, однако не приводят к новым признакам. Среда и условия обитания организма за счет взаимного влияния генов обеспечивают целенаправленное закрепление в потомстве тех признаков, которые более всего им соответствуют.
К этим процессам добавляется еще один генетический фактор. Дело в том, что имеется ряд признаков, непосредственно связанных с полом. Например, в мужской хромосоме Y есть гены, определяющие гемофилию (несвертывание крови), дальтонизм (нечувствительность к красному и зеленому цветам) и ряд других. Эти признаки возникают только у особи данного пола. Надо сказать, что и в этом случае сами гены не меняются, а разнообразие и изменчивость потомства возникает лишь в результате многообразия комбинаций аллелей.
Наряду с этими процессами в природе двуполых организмов заложен процесс изменения признаков - генов половых клеток. Этот процесс именуется мутацией. Мутации могут возникать не только в половых, но и других клетках. Однако они на процесс изменчивости вида никакого влияния не оказывают (если, конечно, организм не размножается в дополнение к половому еще и бесполым путем).
Процесс возникновения мутации в половых клетках именуется генеративным. Причин мутаций известно много: 1) радиоактивные, электромагнитные, химические воздействия на молекулы ДНК, обусловленные нарушением обычных условий среды обитания организма; 2) химическое воздействие различных гормонов, влияющих на работу половых органов; 3) сбои в процессе жизнедеятельности клеток. Большинство мутаций приводят к ослаблению организмов и их потомства. Но некоторые изменения, наоборот, повышают их качественные показатели в борьбе за существование. Именно эти последние постепенно накапливаются и приводят к появлению качественно нового признака. Мутации, приводящие к таким изменениям, носят случайный характер. Однако, судя по результату – постоянному усложнению и усовершенствованию живых организмов, положительные генеративные мутации имеют целенаправленный вектор. У высших животных организмов это связано, по-видимому, с гормональным воздействием на мутации в половых клетках.
В последней трети 20 –го столетия было установлено, что молекула ДНК имеет свойство видоизменяться даже при жизни клетки, в частности, изменяется последовательность нуклеотидных пар, что приводит даже к их увеличению (см. §5.7). Отдельные участки ДНК длиной от тысячи до ста тысяч нуклеотидных пар могут перемещаться вдоль самой молекулы ДНК или переходить на другую молекулу. Такая транспозиция вызывает мутации, обусловленные жизненными процессами, а потому целенаправленно улучшает приспособление организма к условиям среды обитания. Количество таких подвижных пар в ДНК составляет от 10 до 30 %. Сам процесс перемещения явно зависит от условий окружающей среды – если они не меняются, то молекула ДНК не перестраивается, а если происходят изменения, то частота перестройки ДНК половых клеток увеличивается в тысячи и даже десятки тысяч раз.
Ещё одним фактором, играющим важную роль в формировании новых генов, приспосабливающих организм к меняющимся условиям внешней среды, является «мусор»,т.е.совокупность интронов, образующихся в рибосоме в процессе сплайсинга – см. выше [98].
Как и в других природных явлениях, сочетание стохастических и целенаправленных процессов приводят к древовидному накоплению упорядоченности. Отсюда такое многообразие живых организмов и четко выраженный основной ствол развития.
Дата добавления: 2015-08-27; просмотров: 244 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Структура информационной системы клетки | | | Информации |