Читайте также:
|
С точки зрения поведения сон — это период отсутствия активности. Для сна характерно определенное состояние организма, определенный ритм (обычно суточный), повышение порога сенсорных реакций, приуроченность к определенному месту (различные виды убежищ), характерная для данного вида животных поза. Состояние сна у животных может быть определено на основании поведенческих или электрофизиологических критериев, а также на сочетании этих показаний.
Изучение поведения во время сна у морских млекопитающих осложняется их водным образом жизни. В литературе однако имеются описания отдыха у разных видов китообразных в их естественном окружении. Известно (Gray, 1887), что китобойные шхуны подходили вплотную к гренландским китам и финвалам, неподвижно лежащим у поверхности воды или очень медленно передвигающимся. Киты, как бы внезапно очнувшись ото сна, быстро уплывали прочь от корабля.
У островных популяций вертящихся дельфинов наблюдали отдых обычно около полудня (Norris, Dohl, 1980). Они заходили в укромные лагуны и заливы, защищенные от ветра высокими берегами и, сгруппировавшись, медленно двигались по кругу. Группа во время отдыха обычно принимает дискоидальную форму в отличие от построения шеренгой во время активной охоты. У отдыхающих дельфинов резко снижается интенсивность звуковой сигнализации и уменьшается их разнообразие (Powell, 1966; Norris, Dohl, 1980). У вертящихся дельфинов во время отдыха прослушивались только отдельные щелчки. Б. и М. Вюрсиг (Wursig, Wursig, 1979) наблюдали, что в период отдыха тусклые дельфины собираются в тесную труппу и медленно движутся по кругу, представляя как бы единый «амебовидный» организм.
Г. Байтсон (Bateson, 1974), изучая группировки двух видов дельфинов — пятнистых (Stenella attenuata) и вертящихся (Delphinus rosiventis) — в неволе, полагает, что ночные групповые связи отличаются от дневных и истинные социальные связи между членами сообщества можно понять только в их ночных группировках. В период ночного отдыха пять дельфинов из семи проводили ночь вместе. Они медленно двигались по кругу, обычно по часовой стрелке. В это время у них наблюдалось синхронное дыхание. Иногда они меняли направление движения, но при этом у них сохранялись тот же строй и синхронность дыхания. Два самца проводили ночь отдельно от центральной группы. Один из них (лидер) плавал поодаль от группы также по кругу, но его дыхательные движения не были синхронны с остальными дельфинами. Второй самец «зависал» у поверхности воды в характерной позе с опущенным вниз хвостом и выставленной наружу спиной. Днем эти дельфины плавали, группируясь по-другому.
Наблюдения за поведением индийских афалин в океанариуме (Tayler, Saayman, 1972) выявили определенную приверженность дельфинов проводить ночь в одном и том же месте бассейна. Две самки отдыхали вместе у кормовой площадки, два самца — в противоположной стороне бассейна. После того как один самец погиб, второй самец на ночь присоединялся к самкам. В период отдыха самки всегда касались грудными плавниками друг друга.
Ночное поведение афалин, по наблюдению В. Флэнигена (Flanigan, 1974), состоит из периодов стереотипного плавания по кругу с коротким (на 20 — 30 с) закрыванием глаз и неподвижного зависания с признаками сна у поверхности воды. Через каждые 0,5 — 2 мин с помощью слабого удара хвоста дельфин всплывает для очередного вдоха.
Дж. Маккормик (McCormick, 1967) описал у афалин и короткоголовых дельфинов сон, во время которого они погружаются на дно бассейна. При этом виде сна дыхательная пауза у дельфинов, удлиняется до 4 мин. Всплывая на поверхность, они вдыхают несколько раз подряд и снова опускаются на дно.
Афалины во время отдыха принимают самые разные позы. Так, один дельфин, отдыхая, обычно «лежал» на спине у поверхности воды, выставив грудные плавники. На непродолжительное время дельфин, поворачивался брюхом вниз, чтобы сделать вдох и снова принимал прежнее положение (Caldwell, Caldwell, 1972al).
Интересный феномен в поведении дельфинов во время отдыха — попеременное открывание и закрывание глаз — было впервые обнаружено Дж. Лилли (Lilly, 1964). Наблюдая за спящими афалинами, он нашел, что дельфины попеременно открывают то один, то другой глаз. Удалось подсчитать, что в течение 24 ч оба глаза у дельфинов закрыты одновременно только 5 мин. Исходя из этого, Дж. Лилли сделал предположение, что дельфины спят унилатерально.
Не всегда можно с уверенностью сказать, судя только по поведению дельфинов, спят они или приняли позу отдыха. Только электрофизиологические исследования мозга дельфинов помогли раскрыть многие принципиальные вопросы, связанные с процессом сна у этих животных. Приоритет этих исследований принадлежит советским ученым Л. М. Мухаметову, А. Я. Супину (1978).
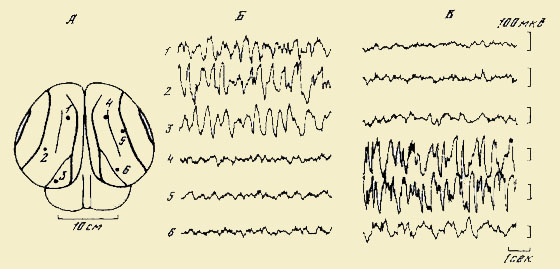
Рис. 16. Межполушарная асимметрия электрокортикограммы спящего дельфина
А — расположение электродов в теменно-затылочной коре; Б, В — одновременно зарегистрированные высокоамплитудные дельта волны в трех точках одного полушария с десинхронизированной электрической активностью симметричных точек коры другого полушария (по: Мухаметов и др., 1976)
Сравнение биоэлектрической активности мозга во время сна у дельфинов и наземных млекопитающих позволило обнаружить некоторые интересные различия (Мухаметов и др., 1976; Мухаметов, Супин, 1978). Во-первых, было выяснено, что дельфины могут спать, находясь в движении; осуществление сложного рефлекторного дыхательного акта не требует их пробуждения. Во-вторых, дельфины могут спать с одним открытым глазом, в то время как у большинства наземных млекопитающих в период сна оба глаза прикрыты веками и предметное зрение выключается. Л. М. Мухаметов и А. Я. Супин показали, что открытый глаз у дельфинов выполняет сложную сторожевую функцию во время сна. Так, внесенный бесшумно предмет в поле зрения открытого глаза сразу вызывал пробуждение дельфина. Авторы полагают, что контроль за изменением среды обитания во время сна у дельфинов осуществляется в отличие от наземных млекопитающих не только слуховым анализатором, к этому контролю присоединяется и зрительная система. В-третьих, у дельфинов была обнаружена межполушарная асимметрия функциональных состояний двух полушарий мозга во время сна, что подтвердило в некоторой степени предположение Дж. Лилли (Lilly, 1964). Во время сна у дельфинов возникает в корковых полушариях поочередно то синхронизация биоэлектрических ритмов (медленные волны), то десинхронизация (ритмика, характерная для бодрствования или парадоксального сна (рис. 16). В этом случае полушария работают как антагонисты: если в левом полушарии записываются медленные синхронизированные волны, то в правом — десинхронизированные, и наоборот. Подобная асимметрия функционального состояния полушарий во время сна не была до сих пор описана у других животных и человека. Смена фаз бодрствование — сон у всех ранее изученных млекопитающих происходит одновременно в обоих полушариях.
Несмотря на то что еще не раскрыты тонкие механизмы, лежащие в основе межполушарной асимметрии различных функциональных состояний мозга дельфинов в период сна, можно полагать, что это явление связано либо с выполнением сторожевых функций одним полушарием, либо имеет отношение к обеспечению дыхательного акта у спящих дельфинов.
Заключение
Жизнь сообществами служит одним из важнейших факторов в борьбе за существование, так как дает огромные преимущества для выживания наибольшего числа потомков. Формы общественного поведения животных, в том числе и у китообразных, в конечном счете направлены на сохранение собственного генотипа в последующих поколениях. Таким образом, стратегия поведения каждой особи в сообществах должна быть направлена на то, чтобы оставить после себя наибольшее число жизнеспособных, плодовитых потомков. Эта цель может достигаться с помощью различных форм социального поведения. Одна из них — установление конкурентных отношений между самцами за преобладающий доступ к самке на основе доминирующей иерархии.
Рассматривая с этой точки зрения групповые взаимоотношения у дельфинов в неволе, можно считать, что у них не существует, видимо, жесткой и стабильной конкуренции за обладание самкой.
Наиболее характерной формой выражения доминирования в сообществах зубатых китообразных можно считать лидерство самцов и самок. Роль лидеров-доминантов в сообществах сводится не только к достижению большего доступа к половым партнерам как средству увеличения числа своих потомков, а и к созданию условий, обеспечивающих их выживание.
В сообществах китообразных детеныши в течение длительного времени окружены заботой как со стороны своей матери, так и других членов сообщества. Длительные и разнообразные формы взаимоотношений матери и ее детенышей в сообществах дельфинов можно рассматривать как важную адаптацию, обеспечивающую выживание их относительно немногочисленного потомства в водной среде, и существенным условием развития и формирования сложных психических функций молодого поколения.
Одной из существенных форм сотрудничества в родственных сообществах зубатых китов является забота о чужих детенышах (аллопарентальное поведение). С эволюционно-генетической точки зрения эта форма поведения рассматривается как способ повышения «приспособленности родственников», которая выражается относительным вкладом в генофонд следующего поколения генов ближайших родственников данной особи.
Взаимопомощь среди взрослых, по преимуществу родственных особей, часто принимающая форму альтруистического поведения, следует также рассматривать как существенный фактор, обеспечивающий выживание этих животных в водной среде. Эти формы общественного поведения, возникшие в родственных сообществах в результате группового отбора, видимо, не следует сводить к простой автоматизированной, видоспецифической реакции. Оказание помощи у дельфинов — весьма лабильная форма поведения, распространяемая не только на особей своего вида, но и на животных других таксономических групп. Лабильность и многообразие проявлений взаимопомощи и альтруистических актов поведения у дельфинов можно связывать с высоким уровнем развития их рассудочной деятельности.
Значение рассудочной деятельности в усложнении организации сообществ и групповых отношений у высших млекопитающих (хищных, приматов и дельфинов) широко обсуждается в литературе (Крушинский, 1977; Hemmer, 1978). При сопоставлении индексов мозга и особенностей социального поведения у разных видов приматов и хищников видно, что относительное увеличение мозга животных коррелирует с пластичностью и сложностью их социального поведения.
У дельфинов, обладающих одним из наиболее высоких коэффициентов цефализации среди животных как можно судить на основании приведенных в этой главе данных, развиваются сложные и многообразные формы общественного поведения, многие из которых характерны также и для высших приматов.
Яркое проявление социальности дельфинов, развитой исследовательской деятельности и способности к «манипулированию» предметами можно найти в игровой активности дельфинов. Игровая деятельность используется дельфинами как средство к установлению контактов с особями своего вида и с животными других видов, а также с человеком. Активное общение с человеком наблюдается у дельфинов не только в условиях неволи как компенсация недостающей естественной активности, но и в природе, где дельфины проявляют инициативу в установлении таких связей с человеком.
Сотрудничество дельфинов с людьми свидетельствует о необычайно гибких и пластичных формах социального поведения дельфинов. Различные виды сотрудничества между человеком и дельфином в природе могли сложиться в результате необычайно развитого у китообразных ориентировочно-исследовательского рефлекса, способности к сложным видам обучения и рассудочной деятельности.
Таким образом, можно заключить, что внутригрупповые отношения у китообразных, складывающиеся на фоне выраженного социального инстинкта, формируются не на основе жестко детерминированной иерархии, а при ведущем участии таких форм поведения, как сотрудничество, взаимопомощь и альтруистические акты поведения. Лабильность и многообразие этих форм поведения могут быть обусловлены высоким уровнем их интеллектуальных возможностей. Это, в свою очередь, видимо, способствовало формированию устойчивых, жизнеспособных сообществ, которые имели существенные преимущества при адаптации к сложным условиям жизни в водной среде.
Дата добавления: 2015-08-27; просмотров: 45 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Общение с человеком и другие сложные формы игрового поведения | | | Нижний Тагил |