|
Читайте также: |
Белково-пептидные гормоны
В процессе образования белковых и пептидных гормонов в клетках эндокринных желез происходит образование полипептида, не обладающего гормональной активностью. Но такая молекула в своем составе имеет фрагмент(ы), содержащий(е) аминокислотную последовательность данного гормона. Такая белковая молекула называется пре-про-гормоном и имеет в своем составе (обычно на N-конце) структуру, которая называется лидерной или сигнальной последовательностью (пре-). Эта структура представлена гидрофобными радикалами и нужна для прохождения этой молекулы от рибосом через липидные слои мембран внутрь цистерн эндоплазматического ретикулума (ЭПР). При этом, во время перехода молекулы через мембрану в результате ограниченного протеолиза лидерная (пре-) последовательность отщепляется и внутри ЭПР оказывается прогормон. Затем через систему ЭПР прогормон транспортируется в комплекс Гольджи и здесь заканчивается созревание гормона. Вновь в результате гидролиза под действием специфических протеиназ отщепляется оставшийся (N-концевой) фрагмент (про-участок). Образованная молекула гормона, обладающая специфической биологической активностью поступает в секреторные пузырьки и накапливается до момента секреции.
При синтезе гормонов из числа сложных белков гликопротеинов (например, фолликулостимулирующего (ФСГ) или тиреотропного (ТТГ) гормонов гипофиза) в процессе созревания происходит включение углеводного компонента в структуру гормона.
Может происходить и внерибосомальный синтез. Так синтезируется трипептид тиролиберин (гормон гипоталамуса).
Гормоны - производные аминокислот
Из тирозина синтезируются гормоны мозгового слоя надпочечников адреналин и норадреналин, а также йодсодержащие гормоны щитовидной железы. В ходе синтеза адреналина и норадреналина тирозин подвергается гидроксилированию, декарбоксилированию и метилированию с участием активной формы аминокислоты метионина.
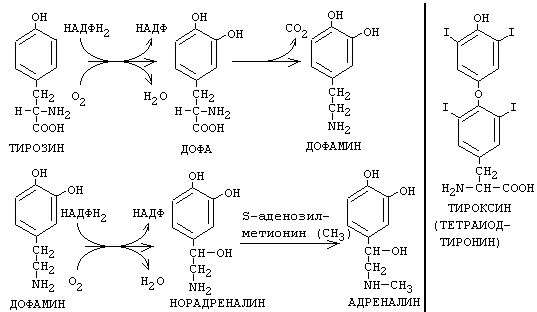
В щитовидной железе происходит синтез йодсодержащих гормонов трийодтиронина и тироксина (тетрайодтиронина). В ходе синтеза происходит йодирование фенольной группы тирозина. Особый интерес представляет метаболизм иода в щитовидной железе. Молекула гликопротеина тиреоглобулина (ТГ) имеет молекулярную массу более 650 кДа. При этом в составе молекулы ТГ около 10% массы - углеводы и до 1% - йод. Это зависит от количества иода в пище. В полипептиде ТГ - 115 остатков тирозина, которые иодируются окисленным с помощью специального фермента - тиреопероксидазы - йодом. Эта реакция называется органификацией йода и происходит в фолликулах щитовидной железы. В результате из остатков тирозина образуются моно- и ди-иодтирозин. Из них примерно 30% остатков в результате конденсации могутпревратитьться в три- и тетра- иодтиронины. Конденсация и иодирование идут с участием одного и того же фермента - тиреопероксидазы. Дальнейшее созревание гормонов щитовидной железы происходит в железистых клетках - ТГ поглощается клетками путем эндоцитоза и образуется вторичная лизосома в результате слияния лизосомы с поглощенным белком ТГ.
Протеолитические ферменты лизосом обеспечивают гидролиз ТГ и образование Т3 и Т4, которые выделяются во внеклеточное пространство. А моно- и дииодтирозин деиодируются с помощью специального фермента деиодиназы и иод повторно может подвергаться органификации. Для синтеза тиреоидных гормонов характерным является механизм торможения секреции по типу отрицательной обратной связи (Т3 и Т4 угнетают выделение ТТГ).
Стероидные гормоны
Стероидные гормоны синтезируются из холестерина (27 углеродных атомов), а холестерин синтезируется из ацетил-КоА.


|
Холестерин превращается в стероидные гормоны в результате следующих реакций:
- отщепление бокового радикала
- образование дополнительных боковых радикалов в результате реакции гидроксилирования с помощью специальных ферментов монооксигеназ (гидроксилаз) - чаще всего в 11-м, 17-м, и 21-м положениях (иногда в 18-м). На первом этапе синтеза стероидных гормонов сначала образуются предшественники (прегненолон и прогестерон), а затем другие гормоны (кортизол, альдостерон, половые гормоны). Из кортикостероидов могут образоваться альдостерон, минералокортикоиды.
СЕКРЕЦИЯ ГОРМОНОВ
Регулируется со стороны ЦНС. Синтезированные гормоны накапливаются в секреторных гранулах. Под действием нервных импульсов или под влиянием сигналов из других эндокринных желез (тропные гормоны) в результате экзоцитоза происходит дегрануляция и выход гормона в кровь.
Механизмы регуляции в целом были представлены в схеме механизма реализации эндокринной функции.
ТРАНСПОРТ ГОРМОНОВ
Транспорт гормонов определяется их растворимостью. Гормоны, имеющие гидрофильную природу (например, белково-пептидные гормоны) обычно транспортируются кровью в свободном виде. Стероидные гормоны, йодсодержащие гормоны щитовидной железы транспортируются в виде комплексов с белками плазмы крови. Это могут быть специфические транспортные белки (транспортные низкомолекулярные глобулины, тироксинсвязывающий белок; транспортирующий кортикостероиды белок транскортин) и неспецифический транспорт (альбумины).
Уже говорилось о том, что концентрация гормонов в кровяном русле очень низка. И может меняться в соответствии с физиологическим состоянием организма. При снижении содержания отдельных гормонов развивается состояние, характеризуемое как гипофункция соответствующей железы. И, наоборот, повышение содержания гормона - это гиперфункция.
Постоянство концентрации гормонов в крови обеспечивается также процессами катаболизма гормонов.
КАТАБОЛИЗМ ГОРМОНОВ
Белково-пептидные гормоны подвергаются протеолизу, распадаются до отдельных аминокислот. Эти аминокислоты вступают дальше в реакции дезаминирования, декарбоксилирования, трансаминирования и распадаются до до конечных продуктов: NH3, CO2 и Н2О.
Гормоны - производные аминокислот подвергаются окислительному дезаминированию и дальнейшему окислению до СО2 и Н2О. Стероидные гормоны распадаются иначе. В организме нет ферментных систем, которые обеспечивали бы их распад. Что же происходит при их катаболизме?
В основном происходит модификация боковых радикалов. Вводятся дополнительные гидроксильные группы. Гормоны становятся более гидрофильными. Образуются молекулы, представляющие собой структуру стерана, у которого в 17-м положении находится кетогруппа. В таком виде продукты катаболизма стероидных половых гормонов выводятся с мочой и называются 17-КЕТОСТЕРОИДЫ. Определение их количества в моче и крови показывает содержание в организме половых гормонов.
Секреция гормонов. Экзоцитоз, эндоцитоз, диффузия. Типы секреции: тоническая и циклическая. Суточные и сезонные ритмы секреции гормонов. Регуляция синтеза и секреции гормонов: гипоталамическая, гипофизарная, метаболическая. Ауторегуляция. Принцип М.М.Завадовского «плюс-минус взаимодействия в эндокринной системе».
Гормональная регуляция, регуляция жизнедеятельности организма животных и человека, осуществляемая при участии поступающих в кровь гормонов; одна из систем саморегуляции функций, тесно связанная с нервной и гуморальной системами регуляции и координации функций.
Одним из важнейших биологических процессов является регуляция секреции гормонов, обеспечивающая их образование, выделение из клеток и поступление в циркуляцию в количестве, необходимом для поддержания процессов метаболизма и других функций тканей и органов. Составными частями этой регулирующей системы являются гуморальные факторы, к которым надо отнести продукты метаболизма и гормоны, нейро-гормональные и нервные факторы.
Можно привести ряд примеров влияния продуктов метаболизма на различные этапы секреции гормонов. Так, примером гуморальных регуляций является выделение инсулина из бета-клеток островков поджелудочной железы во внеклеточное пространство и циркуляцию, при повышении уровня гликемии, тимуляторами этой секреции являются также аминокислоты, оординированно с процессом выделения инсулина происходит повышение его биосинтеза. Снижение уровня сахара крови способствует понижению секреции инсулина, повышению секреции и поступлению в циркуляцию его гормональных антагонистов — глюкагона, вырабатываемого альфа-клетками островков поджелудочной железы, гормона роста, гидрокортизона, адреналина и медиатора норадреналина. Это строго координированное взаимодействие ряда гормонов в итоге сложных метаболических процессов обеспечивает сохранение физиологического уровня сахара крови и метаболизма глюкозы.
Кроме регуляции секреции гормонов в ответ на повышенный к ним запрос, существенное значение имеет высвобождение гормонов из их связи с белками. Изучены специфические белки, связывающие в плазме крови инсулин, тироксин, гормон роста, прогестерон, гидрокортизон, кортикостероп и другие гормоны. Гормоны и протеины связаны нековалентными связями, обладающими сравнительно низкой энергией, поэтому эти комплексы легко разрушаются, освобождая гормон. Комплексирование с белками дает возможность сохранять часть гормона в неактивной форме. Кроме того, эта связь защищает гормон от действия химических и энзиматических факторов. К представлению, что связанные с белками гормоны являются одной из транспортных форм в циркуляции и обеспечивают их резервирование, добавились другие факты: важным компонентом биологического значения этих комплексов является возможность быстрого высвобождения из них свободных, т. е. активных, гормонов.
Регуляция секреции гормонов осуществляется несколькими связанными между собой механизмами. Их можно проиллюстрировать на примере кортизола, основного глюкокортикоидного гормона надпочечников. Его продукция регулируется по механизму обратной связи, который действует на уровне гипоталамуса. Когда в крови снижается уровень кортизола, гипоталамус секретирует кортиколиберин – фактор, стимулирующий секрецию гипофизом кортикотропина (АКТГ). Повышение уровня АКТГ, в свою очередь, стимулирует секрецию кортизола в надпочечниках, и в результате содержание кортизола в крови возрастает. Повышенный уровень кортизола подавляет затем по механизму обратной связи выделение кортиколиберина – и содержание кортизола в крови снова снижается. Секреция кортизола регулируется не только механизмом обратной связи. Так, например, стресс вызывает освобождение кортиколиберина, а соответственно и всю серию реакций, повышающих секрецию кортизола. Кроме того, секреция кортизола подчиняется суточному ритму; она очень высока при пробуждении, но постепенно снижается до минимального уровня во время сна. К механизмам контроля относится также скорость метаболизма гормона и утраты им активности. Аналогичные системы регуляции действуют и в отношении других гормонов.
 Самое важное значение имеет в регуляции секреции гормонов центральная нервная система. Одной из важнейщих областей ЦНС, координирующей и контролирующей функции эндокринных желез, является гипоталамус, где локализуются нейросекреторные ядра и центры, принимающие участие в регуляции синтеза и секреции гормонов аденогипофиза. Гипоталамо-гипофизарная регуляция осуществляется механизмами, функционирующими по принципу обратной связи, в которых четко выделяются различные уровни взаимодействия.
Самое важное значение имеет в регуляции секреции гормонов центральная нервная система. Одной из важнейщих областей ЦНС, координирующей и контролирующей функции эндокринных желез, является гипоталамус, где локализуются нейросекреторные ядра и центры, принимающие участие в регуляции синтеза и секреции гормонов аденогипофиза. Гипоталамо-гипофизарная регуляция осуществляется механизмами, функционирующими по принципу обратной связи, в которых четко выделяются различные уровни взаимодействия.
Под “длинной” цепью обратной связи подразумевается взаимодействие периферической эндокринной железы с гипофизарными и гипоталамическими центрами (не исключено, что и с супрагипоталамическими и другими областями ЦНС) посредством влияния на указанные центры изменяющейся концентрации гормонов в циркулирующей крови.
Под “короткой” цепью обратной связи понимают такое взаимодействие, когда повышение гипофизарного тропного гормона (например, АКТГ) модулирует и модифицирует секрецию и высвобождение гипофизотропного гормона (в данном случае кортиколиберина).
“Ультракороткая” цепь обратной связи – вид взаимодействия в пределах гипоталамуса, когда высвобождение одного гипофизотропного гормона влияет на процессы секреции и высвобождения другого гипофизотропного гормона. Этот вид обратной связи имеет место в любой эндокринной железе. Так, высвобождение окситоцина или вазопрессина через аксоны этих нейронов и посредством межклеточных взаимодействий (от клетки к клетке) модифицирует активность нейронов, продуцирующих эти гормоны. Другой пример, высвобождение пролактина и его диффузия в межваскулярные пространства приводит к влиянию на соседние лактотрофы с последующим угнетением секреции пролактина.
“Длинная” и “короткая” цепи обратной связи функционируют как системы “закрытого” типа, т.е. являются саморегулирующими системами. Однако они отвечают на внутренние и внешние сигналы, изменяя на короткое время принцип саморегуляции (например, при стрессе и др.). Наряду с этим на указанные системы влияют механизмы, поддерживающие биологический циркадный ритм, связанный со сменой дня и ночи. Циркадный ритм представляет собой компонент системы, регулирующий гомеостаз организма и позволяющий адаптироваться к изменяющимся условиям внешней среды. Информация о ритме день-ночь передается в ЦНС с сетчатки глаза на супрахиазматические ядра, которые вместе с эпифизом образуют центральный циркадный механизм – ”биологические часы”. Помимо механизма день-ночь, в деятельности этих “часов” принимают участие другие регуляторы (изменение температуры тела, состояние отдыха, сна и др.)
Связывание гормонов: специфическое и неспецифическое. Транспортные белки. Физиологическое значение связывания гормонов.
Дата добавления: 2015-08-17; просмотров: 152 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Изучаем коктейльную карту | | | Биохимия гормонов |