|
Читайте также: |
Полости наружного слухового прохода и среднего уха (структуры звукопроводящего аппарата органа слуха) заполнены воздухом, а костный и перепончатый лабиринты внутреннего уха, где находится звуковоспринимающий аппарат уха, – жидкостью. С мембраной овального окна контактирует стремечко (его основание), которое передает на нее колебания системы слуховых косточек, возникающие в результате колебаний барабанной перепонки под действием звуковых волн. Причем основание стремечка не укреплено неподвижно в овальном окне внутреннего уха, а скорее перемещается в нем как поршень в цилиндре, в результате чего колебания цепочки слуховых косточек передаются перилимфе преддверия внутреннего уха, а затем и перилимфе заполняющей костный лабиринт улитки (в первую очередь, перелимфе вестибулярной лестницы, берущей начало от овального окна костного преддверия). Колебания же перилимфы в вестибулярной лестнице через тонкую вестибулярную мембрану (стенку перепончатого лабиринта) вызывают движение эндолимфы в улитковом ходе (перепончатом лабиринте улитки, где заложены слуховые рецепторные клетки). Движение же эндолимфы в перепончатом лабиринте улитки приводит в колебательное состояние его нижнюю толстую стенку (базилярную мембрану), что, с одной стороны, сопровождается контактированием волосковых клеток с нависающей над ними текториальной мембраной, а, с другой – возникновением колебаний в перилимфе барабанной лестницы, контактирующей с округлым окном костного преддверия. В связи с тем, что жидкость – несжимаема, каждый раз, получив толчок в овальном окне, она должна передать его куда-нибудь в другое место. Мембрана округлого окна и служит таким местом, поскольку, обладая эластичностью, она обеспечивает гашение колебаний жидкости в преддверии и улитке (перилимфы в костном лабиринте преддверия и улитки и, как следствие эндолимфы в перепончатом лабиринте улитки), вызванных колебаниями мембраны овального окна. Отмеченное делает возможным нормальное функционирование звуковоспринимающего аппарата внутреннего уха.
Соприкосновение волосков слуховых рецепторных клеток с нависающей над ними покровной (векториальной) мембраной в момент колебания базилярной мембраны перепончатого лабиринта приводит к деформации волосков и возбуждению волосковых клеток, которое передается затем на дендриты слуховых нейронов. Следовательно, слуховые рецепторы в функциональном плане представляют собой вторичночувствующие механорецепторы. Необходимо отметить, что для волосковых клеток в покое характерна довольно большая величина мембранного потенциала, составляющая 160мВ (-80мВ – заряд внутренней поверхности мембраны волосковой клетки и 80мВ – заряд эндолимфы), которая облегчает восприятие слабых звуковых колебаний.
Периферическая часть вестибулярного анализатора
 Периферическая (рецепторная) часть вестибулярного анализатора заложена в перепончатом лабиринте преддверия (маточке и мешочке) и ампул полукружных каналов внутреннего уха. В маточке и мешочке сенсорные образования имеют форму пятна (макулы), над которым нависает желатинозная отолитовая мембрана, включающая кристаллы бикарбоната кальция (удельный ее вес в 3 раза больше такового эндолимфы, заполняющей полость перепончатого лабиринта внутреннего уха). В ампулообразных расширениях перепончатых полукружных каналов рецепторные структуры вестибулярного анализатора имеют вид поперечного гребня (кристы), над поверхностью которого расположена желатинозная мембрана – купула, лишенная включений бикарбоната кальция (в связи с чем ее удельный вес такой же, как и эндолимфы, заполняющей перепончатый лабиринт полукружных каналов). Причем если отолитовая мембрана лежит над пятнами маточки и мешочка наподобие плоского камня над пластиной, то купула, подобно створчатому клапану или шарнирной дверце идет от гребешка прямо поперек ампулы, замыкая ее просвет.
Периферическая (рецепторная) часть вестибулярного анализатора заложена в перепончатом лабиринте преддверия (маточке и мешочке) и ампул полукружных каналов внутреннего уха. В маточке и мешочке сенсорные образования имеют форму пятна (макулы), над которым нависает желатинозная отолитовая мембрана, включающая кристаллы бикарбоната кальция (удельный ее вес в 3 раза больше такового эндолимфы, заполняющей полость перепончатого лабиринта внутреннего уха). В ампулообразных расширениях перепончатых полукружных каналов рецепторные структуры вестибулярного анализатора имеют вид поперечного гребня (кристы), над поверхностью которого расположена желатинозная мембрана – купула, лишенная включений бикарбоната кальция (в связи с чем ее удельный вес такой же, как и эндолимфы, заполняющей перепончатый лабиринт полукружных каналов). Причем если отолитовая мембрана лежит над пятнами маточки и мешочка наподобие плоского камня над пластиной, то купула, подобно створчатому клапану или шарнирной дверце идет от гребешка прямо поперек ампулы, замыкая ее просвет.
Рис. 10. Вестибулярный аппарат (поперечный разрез через пятно эллиптического мешочка и ампулу полукружного канала)
Макулы (пятна) маточки и мешочка преддверия и гребни (кристы) ампул перепончатых полукружных каналов состоят из утолщенного эпителия, включающего волосковые (рецепторные) и поддерживающие (опорные) клетки. Волосковые клетки макул перепончатого лабиринта преддверия воспринимают действие силы тяжести и линейных ускорений на организм. Если голова, а следовательно, и макула наклонены относительно силы тяжести, то отолитовая мембрана также стремится скользнуть наклонно, при этом тянет волоски волосковых клеток, деформация которых приводит к возбуждению волосковых клеток. Волосковые клетки гребней ампул перепончатых полукружных каналов выявляют изменения углового ускорения. Когда голове придано угловое ускорение, эндолимфа в определенном полукружгом канале стремится сохранить неподвижность в силу инерции, а перепончатые стенки движутся относительно ее. Это движение смещает купулу из ее

 |
Рис. 11. Схема строения макулы мешочка и гребня полукружного канала
обычного положения относительно гребешка, смещенная же купула тянет волоски волосковых клеток, деформация которых приводит к возбуждению волоковых клеток.
Волосковые клетки макул маточки и мешочка и гребней ампулообразных расширений полукружных каналов вступают в синаптический контакт с дендритами вестибулярных нейронов, тела которых формируют вестибулярный ганглий, заложенный в толще височной кости. Аксоны же вестибулярных нейронов образуют преддверную (вестибулярную) часть преддверно-улиткового нерва, направляющегося в центральную нервную систему.
Следовательно, вестибулярные рецепторы представляют собой в функциональном плане вторичночувствующие механорецепторы.
Волосковые вестибулярные клетки имеют эпителиальную природу и в зависимости от своей формы бывают двух типов: цилиндрические (столбчатые) и колбовидные (грушевидные). На апикальной поверхности этих клеток расположены стереоцилии (до 80 штук, производные мембраны, называемые также волосками или микроворсинками) и, как правило, одна ресничка – киноцилия. Базальная поверхность волосковых клеток принимает участие в образование синаптического контакта с дендритами вестибулярных нейронов.

Рис. 12. Схема ультрамикроскопического строения вестибулярных рецепторов
Периферическая часть вкусового анализатора
Периферическая часть вкусового анализатора локализована в эпителиальном пласте слизистой начальных отделов пищеварительной трубки и представлена вкусовыми рецепторными клетками, входящими в состав вкусовых почек или луковиц. Вкусовые почкизаложены в эпителии боковых поверхностей сосочков языка (кроме нитевидных), слизистой мягкого неба, миндалин, задней части глотки и верхней поверхности надгортанника. Причем апикальная поверхность луковиц достигает поверхности эпителиального пласта. Общее их количество у человека составляет около 10000.

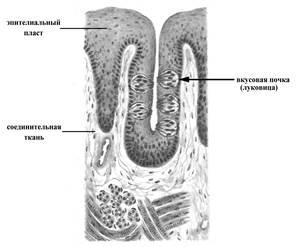
Рис. 13. Листовидные сосочки языка Рис. 14. Схема ультрастуктурной организации вкусовой почки
Вкусовые луковицы имеют колбовидную форму, на их вершине, обращенной к поверхности эпителиального пласта, находится вкусовая пора – отверстие, через посредство которого молекулы различных веществ поступают внутрь почки. Диаметр вкусовых луковиц составляет в среднем 70мкм, они включают два типа клеток: вкусовые (рецепторные) и поддерживающие (опорные). В каждой луковице содержится 2-6 рецепторных клеток, на апикальной поверхности которых имеются микроворсинки (диаметр 0,1-0,2мкм, длина 1-2мкм), увеличивающие площадь поверхности клетки, а следовательно, и площадь ее контакта с молекулами веществ, поступающих в луковицу. Более того, предполагают, что именно в области микроворсинок расположены активные центры – стериоспецифические участки рецептора, избирательно воспринимающие адсорбцию разных веществ и играющие определяющую роль в процессе возбуждения вкусовой рецепторной клетки. Вкусовые клетки характеризуются очень высокой скоростью физиологической регенерации: средняя продолжительность жизни вкусовых эпителиоцитов составляет 250ч (около 10суток), после чего они сменяются молодыми, движущимися к центру вкусовой почки от ее периферии. Базальная поверхность рецепторных вкусовых клеток вступает в синаптичесий контакт с дендритами чувствительных нейронов, тела которых могут быть заложены в чувствительных узлах лицевого (коленчатый узел), языкоглоточного (каменистый узел) или блуждающего (яремный узел) нервов. Аксоны чувствительных нейронов в составе чувствительных ветвей указанных нервов направляются в центральную нервную систему. Следовательно, вкусовые рецепторы являются вторичночувствующими хеморецепторами. С функциональной точки зрения различают 4 типа вкусовых рецепторов, проявляющих наибольшую чувствительность к определенным вкусовым качествам: горькому, соленому, сладкому или кислому. Различные типы вкусовых рецепторов неравномерно распределены на поверхности языка. Так, наибольшая плотность рецепторов, проявляющих максимальную чувствительность к кислому, характерна для латеральных поверхностей языка, сладкому – его кончика, горькому – корня, а рецепторы, наиболее чувствительные к соленому, встречаются по всей поверхности языка.
Периферическая часть обонятельного анализатора
Обонятельный анализатор человека специализируется на восприятии запахов молекул пахучих веществ, попадающих с вдыхаемым воздухом в полость носа и носоглотку. Человек по способности различать запахи относится к микросоматикам (обонятельный анализатор у человека не играет столь важного значения для ориентации в окружающем мире, как, например, у многих ползающих животных (крысы, мыши и другие мелкие грызуны) или у большинства насекомоядных, жвачных, копытных и хищных животных), относящихся к макросоматикам. Его периферическая часть заложена в обонятельном эпителии слизистой носовой полости и представлена собственно дендритами обонятельных нейронов (первичночувствующие хеморецепторы), аксоны которых образуют обонятельный нерв (Iп), направляющийся непосредственно к большому (конечному) мозгу.

Обонятельный эпителий выстилает большую часть крыши каждой полости носа, начинается спереди от переднего конца верхней носовой раковины и простирается назад приблизительно на 1 см; с крыши каждой полости носа он спускается по обеим ее сторонам (причем по медиальной поверхности (носовой перегородке) – приблизительно на 1 см книзу, а по латеральной (крылу носа) вниз почти по всей поверхности носовой раковины).
Рис. 15. Локализация обонятельного эпителия в слизистой носовой полости человека
В обонятельном эпителии различают следующие типы клеток:
Ø собственно обонятельными нейронами, дендриты которых достигают апикальной поверхности эпителиального пласта, на уровне которой заканчиваются обонятельной булавой, от которой отходят 6-12 ресничек (длиной до 10мкм). Считают что в роли обонятельного рецептора, возбуждающегося в результате контакта с молекулами пахучих веществ, выступает обонятельная булава, а отходящие от нее реснички, с одной стороны, увеличивают площадь поверхности булавы, а с другой, могут участвовать в активном захвате молекул пахучих веществ.
Ø малодифференцированными (стволовыми) нейронами, являющимися источником обновления популяции обонятельных нейронов,
Ø поддерживающими (опорными) эпителиоцитами, представляющими собойклетки цилиндрической формы, на апикальном конце которых имеется большое количество микроворсинок увеличивающих площадь их контакта с молекулами пахучих веществ и соответственно концентрацию молекул пахучих веществ в области дендритных разветвлений обонятельных нейронов.
Толщина обонятельного эпителия составляет 100-150мкм, а средний диаметр тел обонятельных нейронов – 5-10мкм. Общее количество обонятельных нейронов у человека достигает около 10млн. В слизистой носа, покрытой обонятельным эпителием, имеются боуменовы железы, секрет которых увлажняет поверхность обонятельных нейронов и поддерживающих эпителиоцитов, способствуя более длительному удержанию молекул пахучих веществ в области обонятельных рецепторов.

Рис. 16. Схема ультраструктур-ной организации обонятельного эпителия
а-дендрит обонятельного ней-рона
б-булавовидная терминаль ден-дрита
в-аксоны обонятельных нейрон-нов, формирующие обонятель-ный нерв, Iп.
г-реснички
Периферическая часть проприоцептивного анализатора
Проприоцептивный анализатор специализируется на восприятии информации о состоянии опорно-двигательного аппарата и косвенно участвует в координации движений.
Периферическая часть проприоцептивного анализатора заложена в опрно-двигательном аппарате (секлетные мышцы, связки, сухожилия, капсулы суставов). Несмотря на существующее разнообразие рецепторов опрно-двигательного аппарата все они с функциональной точки зрения являются первичночувствующими механорецепторами.
Рецепторы скелетных мышц – мышечные веретена (реагируют собственно на изменение длины мышцы, а также на скорость изменения ее длины)
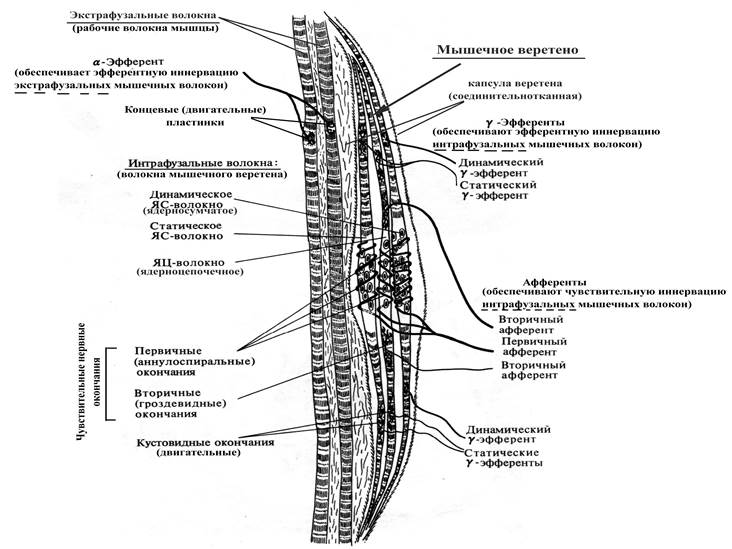
Рис. 17. Схема строения мышечного веретена
Длина мышечных веретен составляет около 3-5 мм, толщина ~0,2 мм, ориентированы в мышцах продольно (в связи с этим при растяжении мышцы растягивается и веретено). Каждое веретено содержит от 2 до 12 интрафузальных мышечных волокон двух типов:
Ø ядерно-сумчатые (в состав веретена входит от 1 до 4 таких волокон, толще и длиннее ядерно-цепочечных, ядра образуют скопления в центре волокна), получают афферентную иннервацию только из одного источника (т.н. первичный афферент, спирально закручиваясь, оплетает ядерную сумку);
Ø ядерно-цепочечные (в составе веретена насчитывается до 10 таких волокон, тоньше и короче ядерно-сумчатых, ядра расположены под сарколеммой в виде цепочки на всем протяжении волокна), получают афферентную иннервацию из двух источников – первичный афферент (более толстый, чем вторичный) оплетает ядерно-цепочечное волокно по центру, а вторичный афферент оплетает ядено-цепочечное волокно по одному с каждой стороны от нервных окончаний первичного афферента.
Интрафузальные мышечные волокна ориентированы параллельно экстрафузальным, в связи с чем при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. Адекватным раздражителем для чувствительных нервных волокон, оплетающих интрафузальные мышечные волокна, является растяжение мышцы, тогда как ее укорочение деактивирует афференты мышечных веретен.
В расслабленной мышце афферентная импульсация от веретен очень слабая, тогда как при растяжении мышцы она резко возрастает. Причем первичные афференты, оплетающие интрафузальные мышечные волокна, реагируют на степень и на скорость растяжения мышцы (динамический ответ), а вторичные, оплетающие ядерно-цепочечные интрафузальные волокна, – только на степень растяжения мышцы (статический ответ).
Наряду с афферентной иннервацией оба типа волокон мышечных веретен получают эфферентную иннервацию со стороны g-мотонейронов. Физиологическая роль эфферентной иннервации интрафузальных мышечных волокон состоит в инициации сокращения их периферических частей, и как следствие, усилении натяжения, а значит и повышении чувствительности на растяжение при любой данной длине мышцы.
Рецепторы связок и сухожилий – тельца Гольджи (реагируют на растяжение связок и сухожилий)
 Рис.18. Схема строения тельца Гольджи
Рис.18. Схема строения тельца Гольджи
Имеют строение, подобное тельцам Руффини соединительной ткани кожи, но отличаются большими размерами. Находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам.
Сухожильные рецепторы Гольджи слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении; интенсивность их импульсации пропорциональна силе сокращения мышцы, что позволяет рассматривать их как источник информации о силе, развиваемой мышцей. На спинальном уровне афферентные нервные волокна, идущие от рецепторов Гольджи, вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антогониста.
Рецепторы капсул сустав – тельца Фатера-Пачини (рецепторы, реагирующие на изменение давления в капсуле сустава, а следовательно, на его положение или изменение суставного угла). Подобно проприорецепторам скелетных мышц и тельцам Гольджи участвуют в системе обратных связей от двигательного аппарата.
Периферическая часть кожного анализатора
Рецепторная часть кожной сенсорной системы заложена в толще кожи (эпителиальном пласте, сосочковом и сетчатом слоях дермы, подкожной жировой клетчатке), площадь поверхности которой составляет от 1,4 до 2,1м2. В функциональном плане чувствительные нервные окончания кожи можно классифицировать на три группы:
Ø тактильные (воспринимают прикосновение, давление, вибрацию)
Ø температурные
ü холодовые (в среднем насчитывается в коже 250тыс., заложены более поверхностно по сравнению с тепловыми (на глубине 0,17мм), максимально возбуждаются при температурах от 10 до 41ºС)
ü тепловые (в среднем насчитывается в коже 30 тыс., заложены более глубоко по сравнению с холодовыми (на глубине 0,3мм), максимально возбуждаются при температурах от 20 до 50ºС)
Ø болевые (ноцицепторы).
В морфологическом плане рецепторы кожи отличаются большим разнообразием, локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. Кожные рецепторы могут быть представлены следующими морфологическими типами:
Ø Свободные нервные окончания (разветвления дендритов чувствительных нейронов). Типичны для эпителиального пласта кожи и роговицы глаза, где могут выступать в качестве механорецепторов (тактильные рецепторы и ноцицепторы) и терморецепторов.
Ø Свободные нервные окончания, оплетающие волосяные луковицы. Типичны для соединительной ткани кожи, имеющей волосяной покров, выступают в качестве механорецепторов (тактильных рецепторов, реагирующих на прикосновение и давление ).
Ø Диски Меркеля. Встречаются в эпителии кожи, причем, как правило, не имеющей волосяного покрова; залегают в глубоких слоях эпителиального пласта (на границе его с соединительной тканью). В функциональном плане диски Меркеля являются механорецепторами, реагирующими на прикосновение и давление (тактильные рецепторы кожи).
Ø Несвободные нервные окончания, лишенные соединительнотканной капсулы. Типичны для соединительной ткани (как правило, рыхлой волокнистой) кожи, где разветвляются параллельно ее поверхности. В функциональном плане могут выступать в качестве механорецепторов (некоторые тактильные рецепторы кожи), терморецепторов и ноцицепторов.
Ø Инкапсулированные нервные окончания (имеют собственную соединительно тканную капсулу)
§ Колбы Краузе. Типичны для соединительной ткани кожных покровов наружных половых органов. В функциональном плане, вероятнее всего, являются первичночувствующими механорецепторами.
§ Тельца Руффини. Типичны для соединительной ткани кожи, особенно, подошвенной поверхности стопы (залегают в сетчатом слое дермы и подкожной клетчатке). В функциональном плане, вероятнее всего, являются первичночувствующими механорецепторами.
Тельца Мейснера. Типичны для соединительной ткани кожи, особенно не имеющей волосяного покрова (пальцы, ладонная поверхность кисти и подошвенная поверхность стопы, губы, веки, наружные половые органы, соски молочных желез). Залегают поверхностно (на границе сосочкового слоя дермы и эпителиального пласта), причем своей длинной осью ориентированы перпендикулярно поверхности кожи; являются первичночувствующими механорецепторами.
§ Тельца Фатера – Пачини. Типичны для соединительной ткани кожи, особенно не имеющей волосяного покрова (пальцы, ладонная поверхность кисти и подошвенная поверхность стопы), а также кожи наружных половых органов и молочных желез. В коже залегают глубоко в дерме (в сетчатом слое, а также в подкожной клетчатке), причем своей продольной осью ориентированы перпендикулярно поверхности кожного покрова. В функциональном плане являются первичночувствующими механо-рецепторами. В коже тельца Фатера-Пачини воспринимают прикосновение и вибрацию.
Следовательно, большинство рецепторов кожи является первичночувствующими.
Дата добавления: 2015-08-02; просмотров: 314 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Взаиморасположение костного и перепончатого лабиринтов улитки внутреннего уха | | | Периферическая часть висцероцептивного анализатора |