Читайте также:
|
Когда ткань возбуждается – генерирует ПД, то временно (соответственно с длительностью ПД) в ней меняется возбудимость: вначале ткань становится совершенно невозбудимой (абсолютная рефрактерность) – любой по силе стимул не способен вызвать в ней новый приступ возбуждения. Эта фаза обычно наблюдается во время пика ПД. Затем происходит постепенное восстановление возбудимости до исходного состояния (фаза относительной рефрактерности) – в этот момент раздражитель может вызвать возбуждение (генерацию нового ПД), но для этого он должен быть намного больше порогового (исходного). Затем (в фазу следовой негативности) возбудимость повышается (супервозбудимость, или фаза экзальтации). В этот момент подпороговые раздражители могут вызывать возбуждение. Наконец, в тканях, в которых ярко проявляется следовая гиперполяризация, наблюдается еще одна фаза – субнормальной возбудимости (сниженной возбудимости).
Биоэлектрические явления в мышцах и нервах. Ток покоя, механизм его возникновения.
Электричество у рыб человек обнаружил еще в глубокой древности. Например, древние греки остерегались встречаться в воде с рыбой, которая, как писал Аристотель, "заставляет цепенеть животных". Рыба, наводившая страх на людей, была электрическим скатом и носила имя "Toгрedo". И только двести лет назад ученые начали приоткрывать природу электричества в живых организмах. Первые научные данные о существовании биоэлектричества ("животное электричество") были получены в 3-й четверти 18 в. при изучении природы "удара", наносимого некоторыми рыбами с электрическими органами при защите или нападении. Было обнаружено, что некоторые рыбы (электрический скат, электрический угорь) при охоте используют электрические разряды, оглушая и обездвиживая свою добычу. В 1791—1792 гг. итальянские ученые Л. Гальвани и А. Вольта первые дали научное объяснение явления "животного электричества". Своими, ставшими уже классическими, опытами они достоверно установили факт существования в живом теле электрических явлений. Позже биоэлектричество было обнаружено и в растительных тканях.
С позиций современных представлений о биоэлектрических явлениях ясно, что все процессы жизнедеятельности неразрывно связаны с различными формами биоэлектрических явлений.
Исследование биоэлектричества имеет большое значение для понимания физико-химических и физиологических процессов в живых системах и применяется в клинике с диагностической целью (электрокардиография, электроэнцефалография, электромиография и др.).
Целью настоящей работы является исследование биоэлектрических явлений. Для достижения названной цели необходимо решить следующие задачи:
1) охарактеризовать понятие биоэлектрических явлений;
2) выявить причины возникновения биоэлектрических явлений;
3) определить показатели биоэлектрической активности.
Понятие о биоэлектрических явлениях
Биоэлектрические явления обусловливают возникновение возбуждения и его проведение по нервным волокнам, являются причиной процессов сокращения мышечных волокон скелетных, гладких и сердечных мышц, выделительной функции железистых клеток и т.д. Биоэлектрические явления лежат в основе процессов всасывания в желудочно-кишечном тракте, в основе восприятия вкуса, запаха, в основе деятельности всех анализаторов и т.д. Нет физиологического процесса в живом организме, который в той или иной форме не был бы связан с биоэлектрическими явлениями. Любой живой организм можно представить в виде сложной смеси жидкостей и различных химических соединений. Многие из этих соединений (и поступающие в организм в виде пищи, и выделенные из него в процессе обмена веществ, и промежуточные вещества, образующиеся при обмене веществ) находятся в виде положительно или отрицательно заряженных частиц — ионов. Перераспределение этих ионов и их транспорт, постоянно имеющие место в процессе жизнедеятельности, — вот причина возникновения биоэлектрических явлений.
Выше уже упоминалось об опытах Л. Гальвани и А. Вольта, достоверно установивших факт существования в живом теле электрических явлений. Однако современные представления о природе биоэлектрических явлений зарождаются в конце 19 - начале 20 вв. Ученый Вебер, проводя вслед за И.П. Тишковым исследования электропроводимости тела живого человека, пытался доказать, что тело человека можно рассматривать в качестве соляных растворов или обычных электролитов. Это положение много лет принималось за верное, хотя многие факты противоречат данным Вебера. Первая достаточно строгая гипотеза была выдвинута Чаговцем (1896), который предложил рассматривать их как диффузионные потенциалы, связанные с неравномерным распределением ионов. Основы современных представлений о механизмах генерации биоэлектрических потенциалов были заложены Ю. Бернштейном (1902 - 1912), связавшим их возникновение со свойствами поверхностной мембраны клетки.
В 1940-х гг. венгерский биохимик Альберт Сцент-Дьёрдьи пришел к выводу, что феномен жизни нельзя должным образом объяснить просто наличием каких-то химических веществ: необходимо, чтобы эти вещества находились в определенном электрическом состоянии. Идеи А. Сцент-Дьёрдьи привели к возрождению интереса к биоэлектричеству. Одним из первых результатов новых исследований в этой области стало обнаружение пьезоэлектрических свойств костной ткани.
Изучение ионной проницаемости мембраны гигантских нервных волокон позволило английским физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947-52) сформулировать современную мембранную теорию возбуждения, принимаемую в настоящее время почти всеми электрофизиологами.
К 1960-м гг. наукой было установлено два фундаментальных положения:
1) электрическая активность свойственна не только животным, но и всем другим биологическим объектам;
2) многие формы электрической активности, наблюдаемые у животных, имеют место и у других организмов.
Биоэлектрические потенциалы
Биоэлектрические потенциалы - электрические потенциалы, возникающие в живых клетках и тканях; показатель биоэлектрической активности, определяемой разностью электрических потенциалов между двумя точками живой ткани. Основными видами биоэлектрических потенциалов являются мембранный потенциал (или потенциал покоя), потенциал действия, постсинаптические потенциалы. Другие виды биоэлектрических потенциалов различных органов и тканей (рецепторные, секреторные, потенциалы сердца, головного мозга и др.) являются аналогами или производными вышеперечисленных биоэлектрических потенциалов. Мембранный потенциал (потенциал покоя) регистрируется между наружной и внутренней сторонами мембраны живой клетки. Его наличие обусловлено неравномерным распределением ионов (в первую очередь ионов натрия и калия) между внутренним содержанием клетки (ее цитоплазмой) и окружающей клетку средой. Внутренняя сторона мембраны заряжена отрицательно по отношению к наружной.
Величина мембранного потенциала различна у разных клеток: для нервной клетки она составляет 60—80 мВ, для поперечнополосатых мышечных волокон — 80—90 мВ, для волокон сердечной мышцы — 90—95 мВ.
При уменьшении потенциала покоя до определенной критической величины (порог возбуждения) возникает кратковременное колебание, получившее название потенциала действия. Если потенциал покоя присущ всем живым клеткам без исключения, то потенциал действия характерен в основном для специализированных возбудимых образований, является показателем развития процесса возбуждения. Вслед за потенциалом действия (пиковый потенциал, или спайк) возникает следовая деполяризация мембраны (отрицательный следовой потенциал) и последующая ее гиперполяризация (положительный следовой потенциал). Амплитуда потенциала действия у большинства нервных клеток млекопитающих составляет 100—110 мВ, у скелетных и сердечных мышечных волокон — 110—120 мВ. Длительность потенциалов действия у нервных клеток 1—2 мс, у скелетных мышечных волокон 3—5 мс, у сердечных мышечных волокон — 50—600 мс. Следовые потенциалы по своей длительности намного превышают потенциал действия. Потенциал действия обеспечивает распространение возбуждения от рецепторов к нервным клеткам, от нервных клеток к мышцам, железам, тканям. В мышечном волокне потенциал действия способствует осуществлению цепи физико-химических и ферментативных реакций, лежащих в основе механизма сокращения мышц.
Постсинаптические потенциалы (возбуждающий и тормозящий) возникают на небольших участках клеточной мембраны (постсинаптической мембране), входящих в состав синапса. Величина постсинаптических потенциалов составляет несколько милливольт, длительность — 10—15 мс. Возбуждающий постсинаптический потенциал (ВПСП) связан с деполяризацией клеточной мембраны. При достижении критической точки деполяризации возникает распространяющийся потенциал действия (см рис. ниже).
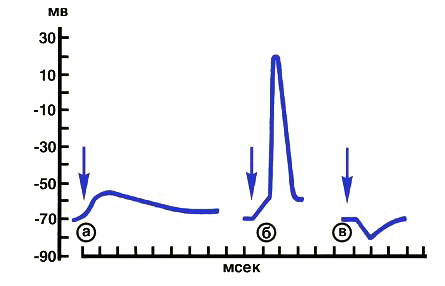
Рис. Схема образования постсинаптических потенциалов: а — возбуждающий постсинаптический потенциал, не достигший величины, достаточной для возникновения потенциала действия; б — возбуждающий постсинаптический потенциал, достигший порогового значения и обусловивший развитие потенциала действия; в — тормозящий постсинаптический потенциал. Стрелки указывают момент действия раздражителя.
Тормозящий постсинаптический потенциал (ТПСП), связанный с гиперполяризацией клеточной мембраны, препятствует возникновению потенциала действия. Механизм возникновения биоэлектрических потенциалов связан с наличием определенных физико-химических градиентов между отдельными тканями организма, между жидкостью, окружающей клетку, и ее цитоплазмой, между отдельными клеточными элементами. Во всех случаях местом возникновения градиентов являются мембраны, различающиеся не только по своей структуре, но и по ионообменным свойствам. Возникновение биоэлектрических потенциалов в живых клетках обусловлено неравномерной концентрацией ионов натрия, калия, кальция и хлора на поверхностях клеточной мембраны и ее различной проницаемостью для них. Величина мембранного потенциала покоя определяется соотношением концентраций, проникающих через мембрану ионов. Высокие концентрационные градиенты ионов калия и натрия поддерживаются благодаря существованию в клеточной мембране так называемого калиево-натриевого насоса, который обеспечивает выделение из цитоплазмы проникающих в нее ионов натрия и введение в цитоплазму ионов К+. Подобный насос работает против их концентрационных градиентов и требует для этого энергии. Источником энергии является аденозинтрифосфорная кислота (АТФ). Энергия, выделяемая при расщеплении локализованной в мембране АТФ-азой одной молекулы АТФ, обеспечивает выделение из клетки трех ионов натрия взамен на два иона калия, поступающих в клетку.
Механизм возникновения потенциала действия обусловлен последовательно изменяющейся во времени проницаемостью мембраны для ионов. Восходящая фаза потенциала действия связана с повышением проницаемости для ионов натрия благодаря все увеличивающемуся количеству открываемых натриевых каналов. Последующая смена активации натриевых каналов на их инактивацию приводит к снижению проницаемости для ионов натрия и возрастанию проницаемости для ионов калия, что приводит к реполяризации мембраны и появлению ее потенциала покоя. В гладких мышцах в отличие от нервных клеток и скелетных мышц в генезе восходящей фазы потенциала действия ведущая роль отводится повышению проницаемости для ионов кальция. В мышце сердца сохранение потенциала действия на определенном уровне (плато потенциала действия) также обусловлено повышением проницаемости мембраны для ионов кальция.
Исследование биоэлектрических потенциалов нашло широкое применение в медико-биологических лабораториях, в клинической практике при диагностике различных заболеваний центральной нервной системы, сердечно-сосудистой и мышечной систем. При отведении суммарных биоэлектрических потенциалов от нервных стволов, мышц, головного мозга, сердца и других органов применяют поверхностные макроэлектроды. В некоторых случаях используют внутриполостные электроды или вводимые непосредственно в ткань (например, игольчатые). Для регистрации и измерения биоэлектрических потенциалов отдельных клеток чаще всего пользуются внутриклеточными и точечно-внеклеточными микроэлектродами. Электроды соединяют с усилителями переменного или постоянного тока, входящими в комплект серийно выпускаемых медицинских приборов. Усилитель может быть связан с устройством автоматизированной обработки биоэлектрических сигналов.
Огромный фактический материал, накопленный к настоящему времени, позволяет говорить о большом разнообразии биоэлектрических потенциалов. В основном они отличаются у разных объектов по двум параметрам:
1) амплитуде
2) частотным характеристикам.
Что касается амплитуды, то разброс величин очень высок. Если разряд электрических рыб (правда, создаваемый не одной клеткой) может достигать 800 вольт, и это достаточно для того, чтобы убить или, по крайней мере, отпугнуть мелкое животное, то, например, регистрируемые с поверхности головы человека биопотенциалы мозга (электроэнцефалограмма) имеют обычно величину порядка нескольких микровольт.
Не менее вариабельны и частотные характеристики биоэлектрических потенциалов. Здесь мы встречаемся, с одной стороны, с практически постоянными во времени величинами. Таковы, например, потенциалы, отводимые от поверхности растений в покое, от поверхности кожи и др.. С другой стороны, имеют место очень быстрые колебания биоэлектрических потенциалов. Так, в серии нервных импульсов позвоночных длительность каждого из них может составлять всего несколько миллисекунд.
Можно различать два типа проявления электрической активности: 1) потенциалы покоя (ПП), то есть разности потенциалов между внутриклеточным содержимым и внешней средой (они регистрируются, когда один микроэлектрод вводится внутрь клетки, а другой находится снаружи поверхностной мембраны; их величина обычно составляет несколько десятков милливольт); 2) потенциалы действия (ПД), то есть изменения ПП при процессе возбуждения.
Итак, на практике все биоэлектрические явления определяют через разность электрических потенциалов между двумя точками живой ткани, которая может быть зарегистрирована специальными электрическими приборами — гальванометрами. С помощью микроэлектродов, например, можно измерить разность потенциалов между наружной и внутренней сторонами оболочки (мембраны) клетки. Эту разность потенциалов называют потенциалом покоя, или мембранным потенциалом.
Биоэлектрические явления в тканях - это разность потенциалов, которая возникает в тканях в процессе нормальной жизнедеятельности.
Живой организм является не только генератором биопотенциалов, но и проводником электрического тока, причем изменение степени электропроводности живых тканей в зависимости от их жизнедеятельности может служить показателем жизнеспособности (состояния) клеток или тканей. Особой формой биоэлектрических явлений служит так называемый электрокинетический потенциал, возникающий, напр., при движении крови по кровеносным сосудам. В этом случае появляется разность потенциалов между стенкой сосуда и движущейся кровью. Величина этого потенциала изменяется при некоторых патологических состояниях, что может быть использовано в диагностических целях.
Возникновением различных форм биоэлектрической активности сопровождается любой акт жизнедеятельности (мышечное сокращение, работа головного мозга, деятельность сердца и т.д.). Регистрация их с помощью специальной аппаратуры расширяет наши возможности в изучении возникновения многих болезней, позволяет диагностировать их. При помощи записи и анализа суммарной электрической активности головного мозга проводят диагностику некоторых нервных и психических заболеваний. На основе изучения электрической активности сердца определяют многие болезни сердечно-сосудистой системы и т.д. На основе биоэлектрических явлений, протекающих в мышцах, созданы биоэлектрические протезы верхних и нижних конечностей.
Тетанус, зубчатый и гладкий. Оптимум и пессимум частоты и силы раздражителя по представлениям Введенского.
Дата добавления: 2015-08-02; просмотров: 98 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Кривая силы-времени. | | | Виды мышечных сокращений |