Читайте также:
|
Еще в XIX веке было установлено, что ультрафиолетовое излучение эффективно уничтожает бактериальные клетки. Однако механизм этого эффекта оставался неясным до тех пор, пока не были измерены его спектры действия. Оказалось, что спектр действия бактерицидного влияния УФ излучения очень близок к спектру поглощения нуклеиновых кислот.
Несколько позже было показано, что к этому же спектру близок и спектр действия мутагенного влияния УФ излучения на проростки кукурузы.
На основании этих и других подобных исследований было сделано заключение о том, что в основе цитотоксического, бактерицидного и мутагенного эффектов УФ излучения лежит поглощение его квантов молекулами нуклеиновых кислот.
Основными хромофорами нуклеиновых кислот, поглощающими в области от 200 до 315 нм, являются пуриновые (аденин и гуанин) и пиримидиновые (цитозин, тимин и урацил) азотистые основания нуклеотидов (рис. 34).
|
|
|
|
|
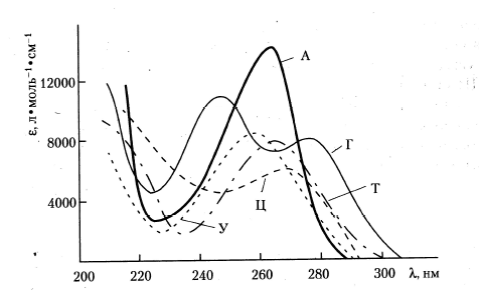
Рисунок 34. Спектры поглощения пуриновых и пиримидиновых оснований нуклеиновых кислот. e - молярный коэффициент поглощения; l - длина волны излучения.
Публикуется с модификациями по: Владимиров Ю.А., Потапенко А.Я. Физико-химические основы фотобиологических процессов. М.: «Дрофа», 2006, с. 120.
Поглощение вызывает преимущественно p®p* переходы, но в длинноволновой части спектра (свыше 280 нм) некоторый вклад вносят также n®p* переходы с участием неразделенной пары электронов атомов азота и кислорода. В растворах при распаде ДНК до смеси свободных нуклеотидов максимум полосы поглощения образца смещается с 258 до 267 нм. Интересно, что оптическая плотность системы при этом увеличивается. Указанные изменения в поглощении носят название гиперхромный эффект. Этот же эффект, хотя и менее выраженный (обычно прирост оптической плотности в этом случае не превышает 40% от ее исходной величины), наблюдается и при денатурации двуспиральной ДНК. Квантовые выходы флуоресценции в водных растворах при комнатной температуре и нейтральных рН у ДНК, РНК и их оснований крайне малы (порядка 10-4). Соответственно малы и времена жизни их возбужденных состояний (порядка 10-12 секунды). Регистрация фотолюминесценции нуклеиновых кислот и их оснований поэтому возможна только при низких температурах и предельных величинах рН среды.
Что касается фотохимических изменений, то следует сразу заметить, что квантовые выходы фотохимических процессов у пиримидиновых оснований намного выше таковых у пуриновых. Поэтому далее основное внимание будет уделено именно фотохимическим реакциям с участием именно пиримидиновых оснований ДНК и РНК.
Пиримидиновые основания, прежде всего, тимин, подвержены фотохимической димеризации:
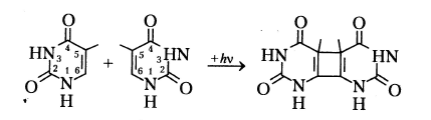
В результате этой реакции формируется циклобутановое кольцо. Именно фотодимеризация тимина вносит наиболее значительный вклад в повреждение ДНК при действии УФ излучения. Например, установлено, что примерно 50% фотоповреждения ДНК клеток кишечной палочки (Escherichia coli) обусловлено именно формированием димеров тимина.
Чаще всего димер формируется между соседними тиминами одной цепи нуклеиновой кислоты. Реже димеризуются расположенные рядом тимины разных цепей. При этом нужно помнить, что в двуспиральной ДНК вследствие принципа комплементарности тимины никогда не располагаются строго друг напротив друга. В то же время, димеризация возможна только тогда, когда тиминовые остатки в разных цепях нуклеиновой кислоты находятся близко друг к другу. Это условие выполняется при локальной денатурации ДНК, при которой двойная спираль нуклеиновой кислоты частично расплетается и позволяет тиминам разных цепей сблизиться. Если при этом под действием излучения происходит их димеризация, формируется зона постоянной денатурации молекулы ДНК, т.е. постоянный дефект этой нуклеиновой кислоты.
Формирующиеся под действием УФ излучения тиминовые димеры весьма устойчивы и не распадаются при нагревании или под влиянием химических факторов. В принципе возможно фотохимическое разрушение этих димеров, но их поглощение лежит в коротковолновой области УФ излучения. Кроме того, если формирование димеров тимина характеризуется спектром действия, соответствующим суммарному спектру поглощения всех оснований нуклеиновой кислоты, разрушение образовавшихся димеров определяется исключительно их собственным поглощением.
Указанные особенности фотодимеризации тимина в молекулах ДНК позволяют добиться того, чтобы в облучаемой нуклеиновой кислоте формировалось определенное количество тиминовых димеров. Если облучение производится средневолновым УФ излучением (l ³280 нм), количество димеров будет нарастать пропорционально дозе облучения. Если же действующее на ДНК УФ излучение содержит кванты с длинами волн около 240 нм, часть формирующихся димеров будет затем подвергнуто фотолизу. Показано, что при воздействии на ДНК УФ излучения при 280 нм в большой дозе до 65% всех тиминов в молекуле нуклеиновой кислоты сформируют димеры. Но при последующем воздействии на такой образец излучением с l =240 нм эти димеры частично будут фотолизированы, и стационарное их количество не будет превышать 17% тиминов облучаемой ДНК.
Поскольку фотодимеризация является бимолекулярной реакцией, ее квантовый выход существенно зависит от среднего расстояния между остатками тимина в нуклеиновой кислоте. Максимальный квантовый выход данной фотохимической реакции наблюдается в политимидиновой кислоте (нуклеиновой кислоте, содержащей только тимины) и составляет примерно 0,02. В случае природных ДНК квантовые выходы фотодимеризации тимина значительно ниже и сильно зависят от воздействий, меняющих пространственную структуру нуклеиновой кислоты (температуры, влажности и т.д.). Последняя особенность позволяет применять оценку эффективности фотоиндуцированного образования димеров тимина в качестве метода исследования пространственной структуры ДНК.
Как уже указывалось выше, спектр действия фотодимеризации тимина в ДНК соответствует суммарному спектру поглощению всех азотистых оснований этой нуклеиновой кислоты. Этот факт позволяет предположить, что реакция фотодимеризации протекает не только за счет поглощения квантов собственно тиминами, но и за счет их поглощения другими основаниями. Вместе с тем, при физиологических условиях азотистые основания отличаются чрезвычайно малыми квантовыми выходами флуоресценции и малыми временами жизни возбужденных состояний. Следовательно, мала и вероятность миграции электронного возбуждения с одного основания на другое по индуктивно-резонансному механизму. Более того, было показано, что индуктивно-резонансная миграция энергии не происходит и при низких температурах, хотя величина квантового выхода флуоресценции и времени жизни возбужденных состояний в таких условиях увеличиваются. Эти факты позволяют предположить, что энергия квантов переносится на тимин с других оснований обменно-резонансным путем, с участием триплетных состояний нуклеотидов. Энергии триплетных состояний мононуклеотидов соответствуют ряду:
Дата добавления: 2015-07-25; просмотров: 3128 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Действие ультрафиолетового излучения на биологические мембраны. | | | Ц > Г > А > Т |