Канонический и альтернативный сплайсинг РНК в клетках человека
Канонический сплайсинг
Механизм сплайсинга пре-мРНК распознает три порции РНК-молекул: 5’-сплайс-сайт, 3’-сплайс-сайт и разветвление интронной последовательности, которое формирует основу выделяемого лариата. Каждая из этих точек имеет универсальную нуклеотидную последовательность, которая остается одинаковой от интрона к интрону и позволяет клетке понять, где начаться сплайсингу.
При сплайсинге ключевые события осуществляются с помощью РНК. Эти молекулы обычно короче 200 нуклеотидов, и 5 из них вовлечены в главные формы пре-мРНК сплайсинга. Они известны как snRNA (от англ. small nuclear RNA). Каждая из них усложняется не менее чем семью белковыми субъединицами, формируя snRNP (small nuclear ribonucleic protein). Эти snRNP формируют основу сплайсосомы.
Сплайсосома – сложный механизм. Многие ученые считают, что она существует в клетке еще до начала процесса сплайсинга. В процессе вырезания интронов, распознавание 5’ и 3’-cплайс-узлов и разветвления интронной последовательности происходит благодаря парности snRNA и универсальных РНК-последовательностей.
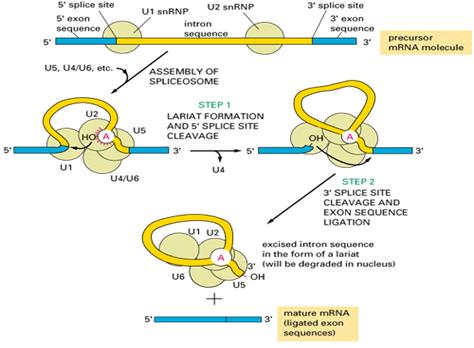
Рисунок 2.1 Схема строения и работы сплайсосомы
(рисунок взят с сайта http://chemistry.umeche.maine.edu/CHY431/Genome2a.html)
Гидролиз АТФ не требуется непосредственно в процессе сплайсинга. Однако он необходим для объединения и преобразования сплайсосомы. Некоторые дополнительные белки используют высвободившуюся энергию для разрушения существующих РНК-РНК связей и образования новых.
АТФ-зависимые РНК-РНК преобразования происходят как внутри snRNP, так и между snRNP и пре-мРНК субстратом. Одна из важнейших их функций – создание активной каталитической точки сплайсосомы. Стратегия образования активной точки только после объединения и преобразования сплайсосомы – эффективный путь предотвращения ошибок.
Каталитическая точка формируется из молекул РНК, а не белков. Благодаря этому, некоторые snRNA формируют трехмерную структуру, которая сопоставляет 5’-конец пре-мРНК с разветвленной интронной последовательностью и производит первую трансэтерификационную реакцию. Подобным образом 3’- и 5’-сплайс-сайты соединяются вместе, осуществляя вторую трансэтерификационную реакцию.
Когда химический акт сплайсинга завершен, snRNP остаются прикрепленными к лариату. Их отделение от него и друг от друга требует еще одной серии РНК-РНК переходов и гидролиза АТФ. Это возвращает snRNA в их первоначальное состояние, и они могут быть использованы снова. После окончания сплайсинга сплайсосома направляет ряд протеинов к мРНК возле места, к которому был прикреплен интрон. Формируя экзонный узловой комплекс, эти белки помечают место успешного акта сплайсинга и влияют на последующую судьбу мРНК.
Интронные последовательности сильно различаются по размеру. Если бы выбор точек сплайсинга обеспечивался только действием snRNP, следовало бы ожидать ошибок, таких, как выпадение экзонов или возникновение неверных сплайс-сайтов. Особые механизмы, встроенные в сплайсосому, дополняются двумя дополнительными стратегиями, которые увеличивают точность ее работы.
Первая – это последовательность ранних стадий сплайсинга, которые происходят еще при синтезе пре-мРНК. В процессе транскрипции фосфорилированный хвост РНК-полимеразы несет компоненты сплайсосомы, которые переносятся точно от фермента к РНК в процессе синтеза. Эта стратегия позволяет сохранить четкое чередование интронов и экзонов, так как snRNP, присоединяющаяся к 5’-сайту сплайсинга, первоначально представлена лишь одним 3’-сайтом сплайсинга, в то время как остальные такие точки ниже по цепи еще не были синтезированы. Координация транскрипции и сплайсинга очень важна для предотвращения нежелательного выпадения экзонов.
Стратегия, называемая «уточнение экзонов», – еще один способ выбора верных точек сплайсинга. Размер экзона считается гораздо более стандартизированным, чем интрона (150 пар нуклеотидов у многих эукариот). В соответствии с этой стратегией, механизм сплайсинга первоначально отыскивает родственные и сходные по размеру кодирующие последовательности. При продолжении синтеза РНК, группа дополнительных компонентов (в основном, SR-белки) присоединяется к экзону и отмечает 5’- и 3’-сайты сплайсинга начиная с 5’-конца РНК. Эти протеины пополняют snRNA U1, которая отмечает границы экзона, и U2AF, которая уточняет вышележащие. Отмечая экзоны и пользуясь преимуществом их одинакового размера, клетка увеличивает точность привнесения начальных сплайс-компонентов возникающей РНК и тем самым позволяет избежать появления неверных мест сплайсинга. Как конкретно SR-белки отличают экзонные последовательности от интронных, пока детально неизвестно. Однако обнаружено, что эти белки прикрепляются преимущественно к особым РНК в экзонах.
Так как несколько разных кодонов могут кодировать определенную аминокислоту, имеется свобода модификации экзонной последовательности таким образом, чтобы сформировать прикрепительную точку для SR-белков без необходимости влияния на аминокислотную последовательность, которая ею определяется.
И мечение границ экзонов и интронов, и соединение сплайсосомы начинается на молекуле РНК, когда она еще удлиняется с помощью РНК-полимеразы на 3’-конце. Однако настоящий химизм сплайсинга может начинаться гораздо позже. Это означает, что интроны не обязательно вырезаются из молекулы пре-мРНК в том же порядке, в котором они располагаются в цепи. (Alberts B. et al, 2007)
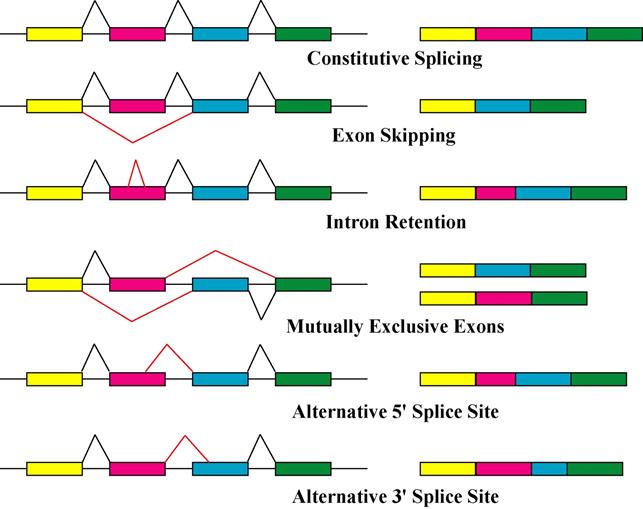
Рисунок 2.2 Другие возможности сплайсинга
(рисунок взят с сайта http://pdflockers.com/pdf/rna-splicing-wikipedia-the-free-encyclopedia)
Дата добавления: 2015-09-07; просмотров: 181 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| РНК в ядре клеток человека | | | Разнообразие, строение и функции |