Читайте также:
|
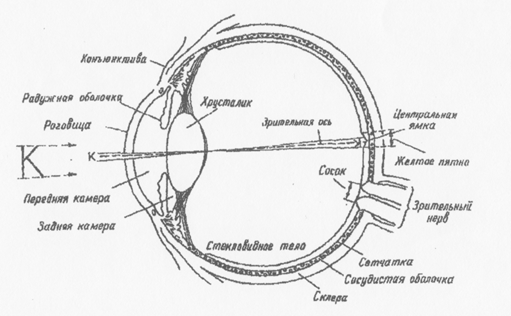
Светочувствительной системой глаза является сетчатка, где
происходит преобразование световой энергии в электрическую, Сетчатка содержит рецепторы, несколько слоев нервных клеток и слой пигментных слеток.
На сетчатке имеется два пятна: желтое пятно или центральная ямка - наиболее чувствительная часть сетчатки и слепое пятно - не имеет рецепторов, это место выхода зрительного нерва (Рис.5).
Рецепторы - это палочки и колбочки (Рис.6).
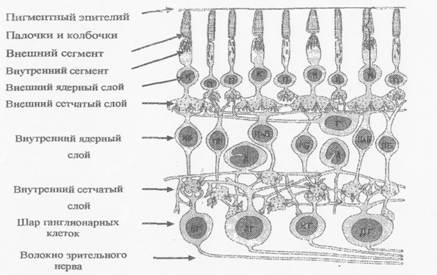
Рис.6. Нейрональные компоненты внеямочной чаеги сетчатки: К - колбочка; П - палочка; КБ, ПБ, ПлБ - карликовые, палочковые и плоские биполярные клетки; ДГ, КГ- диффузные и карликовые ганглионарные клетки, Г - горизонтальные клетки; А - амакриновы клетки
Каждый рецептор состоит из наружного сегмента, который содержит зрительные пигменты, и внутреннего сегмента, который содержит много митохондрий, ядро, аппарат Гольджи и др. органеллы, обеспечивающие энергетический и пластический обмен, в частности синтез белка и АТФ.
Палочки, их около 120 млн., расположены на периферии сетчатки, в центральной ямке полностью отсутствуют. Это рецепторы ночного видения, содержат светочувствительный пигмент родопсин. Палочка в 500 в более чувствительна к свету, но не реагирует на свет с разной длиной волны т.е. она не цветочувствительна. Колбочки, их около 6 млн., росположены, в основном, в центре сетчатки. Это рецепторы дневного дения с меньшей чувствительностью, но участвуют в цветовосприятии, держат зрительные пигменты: иодопсин (желтый), хлоролаб (зеленый), итролаб (красный). Зрительные пигменты обоих рецепторов находятся мембранных дисках внешних сегментов: только в палочках они делены от внешней мембраны своей собственной мембраной, а в лбочках представлены складками плазмолеммы с общей клеточной мебраной (Рис.7).
Рис.7. Схема строения палочек и колбочек

Пигментный слей содержит черный пигмент - меланин. Пигмент, поглощая свет, препятствует его отражению от стенок и попаданию на соседние рецепторы, чем и обеспечивают четкость видения. Пигментный слой содержит большое количество витамина А, участвующего в ресинтезе зрительных пигментов. Эти клетки участвуют в фагоцитозе наружных сегментов палочек, которые обновляются каждые 2-3 недели и колбочек, восстанавливающихся чуть реже, защищают фоторецепторы от повреждений избыточным светом способствуют переносу кислорода и др. необходимых веществ к фоторецепторам. Контакт между клетками пигментного слоя и фоторецепторами довольно слабый, что иногда приводит к отслойке сетчатки, далее к дегенерации рецепторов и потере зрения.
Биполярные клетки - принимают информацию от рецепторов и передают ее через синапсы к танглиозным клеткам.
Ганглиозные клетки - передают информацию в вышележащие структуры.зрительного анализатора, аксоны их формируют зрительный нерв.
Горизонтальные клетки, соединяя несколько рецепторов и биполярную клетку, выполняют функцию латерального торможения на уровне биполярный клеток.
Амакрииовые клетки - возбуждаются от биполярных и выполняют также функцию латерального торможения, но уже на уровне ганглиозных клеток.
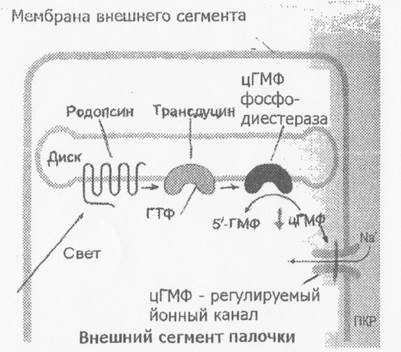
Обработка зрительной информации начинается с возбуждения фоторецепторов. При действии кванта света на зрительные пигменты фоторецепторов в них происходит цепочка фотохимических реакций (Рис.8).
Ри.8. Начальные этапы превращения света в палочках.
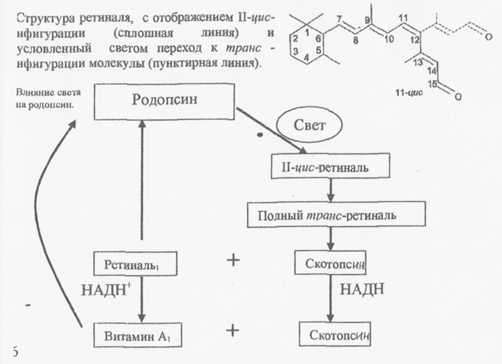
В молекулы зрительного пигмента, находящиеся на диске наружного сегмента палочек и колбочек, состоят из большой белковой части - опсина и меньшей части - ретиналя или альдегида витамина А. Ретиналь может находиться в различных пространственных конфигурациях, т.е. стереоизомерных формах, но только одна из них - II - цис - изомер ретиналя выступает в качестве хромоформной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды, при недостатке которых развивается «куриная слепота» - нарушение зрения в темноте при хорошем видении днем.
При поглощении кванта света в молекуле родопсина идет мгновенная изомеризация цис-формы в трансретиналь. Вслед за фотомеризацией ретиналя происходят пространственные изменения в белковой части - онсине. В результате родопсин обесцвечивается и переходит в состояние метародопсииа II, который способен к взаимодействию с белком трансдуцином (Рис. 8). Трансдуцин обменивает ГДФ на ГТФ, что в свою очередь активирует фосфодиэстеразу (ФДЭ). Активированная ФДЭ разрушает молекулы цГМФ, превращая его в 5-ГМФ. Это приводит к закрытию Na+ каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь рецептора входили и Са2' Ионный канал закрывается вследствие того, что из-за падения концентрации свободного цГМФ в клетке от канала отходят молекулы цГМФ, которые были связаны с ним в темноте и держали его открытым. МП палочки в темноте равен в связи со свободным заходом Ыа - 25мВ. На свету, когда уменьшается вход Na+ МП становится - 90мВ, т.е. наступает гиперполяризация. Т.о. рецепторный потенциал палочек и колбочек связан не с деполяризацией, а с гиперполяризацией. Следовательно с пресинаптического окончания рецептора на биполярную клетку прекращается выделение медиатора (глутамат).
Не менее сложен и механизм восстановления исходного темнового состояния фоторецептора, т.е. его способности ответить на следующий световой стимул. Всё идёт в обратном направлении. Для этого необходимо открыть Na+ каналы. Открытое состояние канала обеспечивается его связью молекулой цГМФ, что в свою очередь регулируется концентрацией свободного цГМФ в цитоплазме, т.е. снижается активность ФДЭ в темноте, значит растет уровень цГМФ в цитоплазме и открываются Na+ каналы. Na+ и Са2+ вновь заходят, деполяризуя мембрану рецептора и переводя его в "темновое" - тормозное состояние. Вследствие этого из пресинаптического окончания вновь ускоряется выход тормозного медиатора в синапсе рецептор - биполяр.
На 130млн фоторецепторов приходится 1,25млн ганглиозных клеток. Это значит, что импульсы от многих ФР сходятся (конвергируют) через молярные нейроны к одной ганглиозной клетке. Фоторецепторы, соединенные с одной биполярной, а дальше и ганглиозной клеткой образуют ее рецептивное поле (РП). Биполярная и ганглиозная клетка, возбуждения на себе повышает световую чувствительность, но ухудшает пространственное разрешение, чем снижается острота зрения, лишь в центре сетчатки (желтое пятно) каждая колбочка соединена с одной биополярной и ганглиозной клеткой. Это обеспечивает здесь высокое пространственное разрешение, но резко уменьшает световую чувствительность.
Также различают биполяры оn-типа: если освещен центр рецептивного поля они деполяризуются (возбуждение), если периферия гиперполяризуються (торможение); и off-типа - при освещении центра гиперполяризуются, а периферии - деполяризуются (Рис.9). 
Рис. 9. Импульсация двух ганглиозных клеток сетчатки и их концентрические рецептивные поля. Тормозные зоны рецептивных полей заштрихованы. Показаны реакции на включение и выключение света при стимуляции световым пятном центра рецептивного поля и его периферии.
Функциональная организация слоя биполярных клеток определяются двумя важнейшими принципами обработки зрительной информации нейронами.
1. Возбуждение фоторецепторов передаётся в ЦЙС по двум независимым «каналам»: через оп-систему. активируемую, когда световая стимуляция в центре сильнее, чем на периферии или чем предыдущая стимуляция центра, и через оff-систему, активируемую при уменьшении освещения центра.
2. РП организованы антагонистически, т.е. свет вызывает их диаметрально противоположную реакцию в зависимости от того, попадает он на периферию или в центр РП. Такая пространственная организация РП обеспечивает конвергенцией сигналов от нескольких рецепторов на одной биполярной клетке и опосредованным торможением с периферии РП через горизонтальные клетки (латеральное торможение).
Принцип конвергенции сигналов и латерального торможения действует на уровне ганглиозных клеток и амакриновых клеток.
Ганглиозные клетки с оnn-центром деполяризуются в ответ на освещение центра РП, и гиперполяризуются в ответ на выключение света в центре или освещение периферии. «Латерально» тормозятся памакриновыми клетками и латерально возбуждаются off- амакриновыми, возбуждаемыми off-биполярами. Если и центр и периферия РП освещаются одновременно, доминирует реакция центра. Ганглиозные клетки с off-центром работают наоборот.
Т.о. на уровне ганглиозных клеток возникают концентрические поля, состоящие из возбужденных и заторможенных участков, т.е. формируется своеобразный «узор» передающий (сканирующий) РП, фоторецепторов, воспринимающих световое раздражение.
По зрительным нервам информация поступает в верхние бугры четверохолмия (средний мозг), затем в латеральное коленчатое тело (таламус) и, наконец, в центральное звено - зрительную кору (затылочные доли), где она декодируется и превращается в конкретное изображение.
МЕХАНИЗМ СВЕТОВОЙ И ТЕМНОВОЙ АДАПТАЦИИ ГЛАЗА
Адаптация глаза – это приспособление глаза к обеспечению остроты зрения на свету и в темноте. Адаптацию обеспечивают процессы, происходящие в сетчатке на уровне рецепторов, а также изменение освещенности сетчатки за счет диаметра зрачка.
Механизмы темновой адаптации:
1)Повышение световой чувствительности палочек. Оно связано с увеличением образования родопсина, т.к. на свету большое количество этого пигмента разрушается. В темноте, когда длительное действие раздражителя (света) становится слабым, необходим усиленный синтез его для нормального функционирования палочек. Глаз переключается на ночное скотопическое зрение. Это длится около часа. Восстановление родопсина сопровождается резким (100-200тыс. раз) повышением чувствительности палочек к свету.
2)Происходит рефлекторное расширение зрачка:
B. для проникновения в глаз большого количества света;
C. для хорошого освещения не только центра, но- и периферии сетчатки, где расположены палочки.
Одновременно с расширением зрачков снижается острота зрения. Острота зрения - это способность глаза различать четко две точки, которые расположены на минимальном расстоянии друг от друга. Иными словами - это наименьшее расстояние, на котором две точки воспринимаются как отдельные, не сливаясь. Остроту зрения определяют по таблицам Сивцева, где есть 10 рядов оптотипов с уменьшающимися размерами. Один ряд соответствует 0,1 единице остроты зрения. Нормальная острота зрения принята за 1 единицу. Испытуемый с нормальной остротой зрения должен с расстояния 5м под углом зрения в 1 различить все 10 рядов оптотипов.
Так как в сумерках, во-первых, подключаются палочки, которые находятся на периферии сетчатки; во-вторых, происходит расширение зрачка, то имеет место сферическая абберация глаза, в результате которой часть лучей фокусируются не строго на сетчатке, а перед или за ней. В результате разницы фокусного расстояния при преломлении по центру и по периферии роговицы и хрусталика изображение получается не очень четким, острота зрения снижается.
Существенную роль в темновой адаптации также играет переключение связей между элементами сетчатки. В темноте площадь рецептивного поля ганглиозной клетки, которая и так достаточно велика из-за периферически расположенных палочек, еще больше увеличивается вследствие ослабления горизонтального торможения горизонтальными и амакриновыми клетками. При этом увеличивается конвергенция фоторецепторов на биполярные нейроны и биполярных нейронов на ганглиозную клетку. Вследствие такой пространственной суммации световая чувствительность на периферии сетчатки в темноте возрастает, а острота зрения падает.
Темновая адаптация заканчивается в течение 20 минут, хотя незначительное снижение порога светового раздражения прослеживается и позже (до 50 мин).
При световой адаптации наоборот:
1) Снижается световая чувствительность колбочек. Во-первыхэто связано с подключением зрительных пигментов, имеющих значительно больший порог раздражения - фотопсинов, т.н. дневных, обеспечивающих ф отоптическое зрение. Во-вторых, на каждую ганглиозную клетку конвергирует одна или несколько колбочек, что снижает степень пространственной суммации, а значит и световую чувствительность. При этом повышается разрешающая способность ганглиозных клеток, увеличивается острота зрения. В - третьих повышается тормозной контроль со стороны коры на сетчатку (эфферентные влияния).
1) Происходит сужение зрачка (5сек.), что также снижает чувствительность колбочек из-за уменьшенного потока света.
2) Повышается острота зрения, т.к. сужение зрачка, резко ограничивая световой поток, направляет его строго в центральную ямку, тем самым способствует исчезновению сферической абберации. Изображение фокусируется строго на сетчатку.
Длительность световой адаптации гораздо меньше - до 5 минут. По сути, световая адаптация сводится к исчезновению темновой адаптации.
Глаз человека способен различать цветовую гамму различных длин волн в диапазоне от 400 до 760 нм. Полагают, что на уровне рецепторов цветовое зрение обеспечивается наличием на сетчатке трех типов колбочек: одни содержат пигмент, реагирующий на красный цвет (эритролаб), другие - на зеленый цвет (хлоролаб), третьи - на синий (иодопсин). Другие цвета возникают при возбуждении неодинаковых количеств этих трех видов колбочек.
Это так называемая трехкомпонентнаи терия цветового зрения, создателями которой являются Ломоносов, Юнг, Гельмгольц.
Существует и другая теория - оппонентных цветов (Геринг). По ней предполагается, что в сетчатке имеется три пары колбочек: 1 воспринимающих красно-зеленый цвет; 2) воспринимающих черно-белое; 3) воспринимающих желто-синий цвет. Так, в сетчатке есть нейроны, активируемые красным и тормозимые зелёным светом в их РП (простые оппонентные клетки). ВКГ и зрительной коре есть ещё более сложные которые дают возбуждение на красный цвет и торможение на пёный еще и с учетом освещения центра и периферии (дважды опонентные клетки). Например, если красный цвет в центре РП збуждает нитроны, а зелёный-тормозит, то на периферии РП картина противоположная.
Обе теории логично обоснованы и взаимодополняют одна другую.)трехкомпонентная теория пригодна для цветовосприятия на уровне лбочек. А обработка цветовой информации на более высоких уровнях, начиная с ганглиозных нейронов, происходит по типу одновременного цветового контраста.
Люди с нормальным цветовосприятием - трихроматы. Дихроматы и трихроматы - это люди, которые имеют лишь две или одну колбочковую систему. Дейтеранопы - не воспринимают зеленый спектр, протанопы - красный, тританопы - синий. Полная цветовая слепота - ахромазия - всегда человек видит все в серых оттенках.
ОБРАБОТКА ЗРИТЕЛЬНОЙ ИНФОРМАЦИИ В ПОДКОРКОВЫХ ЯДРАХ И В КОРЕ ГОЛОВНОГО МОЗГА
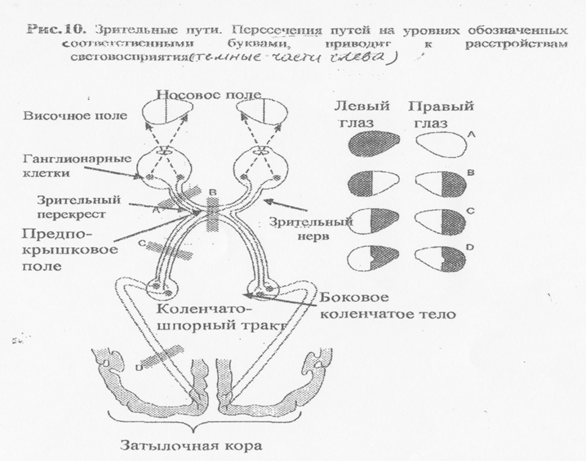
От обеих глаз отходят правый и левый зрительный нервы, которые не прыяиваясь в области хиазмы опять расходятся в соответствующие (правую и левую) части подкорковых ядер (Рис.10). В хиазме происходит скрещивание части волокон, несущих информацию от внутренних частей противоположных участков сетчатки. Т.е. каждый зрительный нерв после хиазмы проводит импульсы частично (от латеральной стороны сетчатки) своей стороны и частично (от медиальной стороны сетчатки) - противоположной. Частичный перекрест волокон обеспечивает каждое полушарие большого мозга информацией от обоих глаз. Проекции эти организованы так, что в затылочную долю правого полушария поступают сигналы от правых половин сетчаток, а в левое - от левых.Т.о. изображение левых половин полей обоих г лаз, преломляясь проецируется в правое полушарие и, наоборот, правые поля - в левое. Зрительные пути достигают среднего мозга и прерываются в верхних буграх четверохолмия. Здесь идет различение движущихся стимулов. С этих подкорковых ядер обеспечивается выполнение двигательных рефлексов, а также саккадических движений глаз. Передние бугры четверохолмия являются первичными центрами интегрирования сенсорной информации, используемой для пространственной ориентации.
В следующем подкорковом центре зрительного проводникового пути - латеральном коленчатом теле таламуса - нейроны сгруппированы в вертикальные колонки одинакового функционального на значения. Мелкоклеточные нейроны проводят возбуждения, передающие форму, цвет, а крупноклеточные - отвечают за различения движений, расположение, пространственную организацию. Как и ганглиозные клетки сетчатки нейрона, ЛКТ имеют концентрически организованные рецептивные поля. Классификация этих клеток примерно такая же, как в сетчатке: часть отвечает на контрастность, часть - на локальные значения света и тени, часть - на цветовосприятия. ЛКТ связано со стволовыми структурами, которые добавляют в обработку зрительной информации степень бодрствования, внимания, движения глаз.
 | ||
 |

Т.о. один из принципов функциональной организаций зрительной коры послойное расположение клеток с рецептивными полями параллельными её поверхности. Как и в других корковых зонах, здесь наблюдается второй принцип - геометрическая организация, перпендикулярная поверхности мозга, т.е колончатое строение. А это значит:
а) рецептивные поля нейронов в пределах «колонки» расположены в одной и той же части сетчатки;
б) нейроны колонки возбуждаются сигналами либо от левого, либо от правого глаза, да еще с учетом ориентации каждого глаза;
с) выделяются цветоспецифические колонки.
В первичной зрительной зоне коры одни нейроны реагируют на просгые стимулы типа свет или темнота, другие - на простые контуры определенной ориентации, их изломы. Сложные нейроны связаны с перцептивными полями, стимулом для которых являются границы между те:мным и светлым или разрывы границ определенной ориентации и ширины и т.д. Сверхсложные РП нейронов реагируют на границы между светлым и темным строго определенной ориентации и ограниченной длины, определенные углы и т.н. Поля 18 и 19 представляют вторичную чувствительную зону коры, которые также учавствуют в обработке зрительной информации. Итак, от уровня к уровню зрительной системы происходит ос:ложнение рецептивных полей нейронов. Все рецептивные поля организованы в виде возбудительных и тормозных зон. Концентрические перцептивные поля характерные для сетчатки и латерального коленчатого тела, уже в коре выглядят прямоугольными.
Время инерции «зрения» в среднем равно 0,03-0,1сек. Это ощущение исчезает также не сразу после того, как прекратилось раздражение, оно еще держится некоторое время и называется последовательным образом, Ес ли посмотреть на лампочку и закрыть глаза, то она видна еще некоторое время (положительный последовательный образ). Если же после рассмотрения лампочки взгляд перевести на светлый фон, то некоторое время держится негативное изображение, т.е. светлые части будут темными, а темные - светлыми (отрицательный негативный образ).Причина его в том, что возбуждение от освещенного объекта тормозит (адаптирует) определенные участки сетчатки; если перевести взор на освещенный экран, то его свет сильнее возбудит те участки, которые не были возбуждены раннее.
ФУНКЦИИ НАРУЖНОГО, СРЕДНЕГО И ВНУТРЕННЕГО УХА
Функциями наружного уха являются:
1) улавливание и проведение звука вдоль наружного слухового прохода до барабанной перепонки;
2) ограничение потока звуковых сигналов с тыльной стороны головы;
3) обладая акустическими свойствами, т.е. имея собственную (резонансную) частоту колебаний равную 3 кГц, обеспечивает минимальный порог слышимости, особенно в речевой области;
4) поддержание постоянства влажности, температуры у барабанной перепонки;
5) защищает слуховой аппарат среднего и внутреннего уха, вырабатывая серу, которая задерживает пылевые частицы;
6) барабанная перепонка воспринимает звуковые колебания и передаст их в среднее ухо.
Функции среднего уха:
1. проведение звука,
2. усиление звука;
3. защитное действие. Проведение звука, который представляет собой колебания молекул воздуха в виде продольной волны давления, осуществляется путем превращения звуковых колебаний в механические колебания косточек (молоточек, наковальня, стремечко) (Рис.12). При этом происходит снижение амплитуды колебаний при одновременном их усилении.
Усиление давления звуковых волн осуществляется благодаря а) наличию системы рычагов, которые увеличивают силу колебаний в 1,3 раза, а также, б) различию поверхностей стремечка и барабанной перепонки: площадь последней в 22 раза больше. Следовательно, давление звуковой волны на овальное окно в 22 раза становится сильнее, чем при ее попадании на барабанную перепонку. Все это создает чрезвычайно высокую чувствительность слухового анализатора, что даже слабые звуковые волны действующие на барабанную перепонку способны преодолеть сопротивление мембраны овального окна преддверия и привести к колебаниям жидкости в улитке. Такое усиление несет большую функциональную роль, т.к. жидкость обладает значительно большим акустическим сопротивлением, чем воздух. Благоприятные условия для колебания барабанной перепонки создает евстахиевая труба. Соединяя среднее ухо с носоглоткой, она служит для выравнивания давления в среднем ухе с атмосферным.
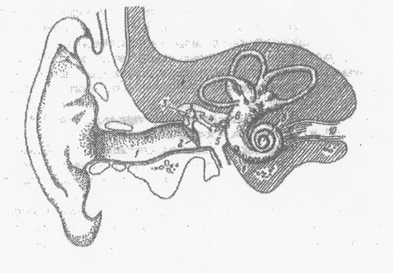
Рис 1. 2. Схема наружного, среднего и внутреннего уха.
1 — слуховой проход, 2 — барабанная препонка, 3 — молоточек, 4 — наковальня, 5 — стремечко,
6 — овальное окно, 7 — полукружные каналы, 8-улитка, 9 — круглое окно, 10 — слуховой нерв.
Защитные действия проявляются в следующем: расположенные в полости среднего уха мышцы, влияя на натяжение барабанной перепонки и ограничивая амплитуду движения стремечка при сокращении, повышают «жесткость» мембран (барабанной перепонки, овального окна), делают их менее подвижными и чувствительными к сильному звуку и приводят соответственно к уменьшению звукового давления во внутреннее ухо. Обеспечивают защитный акустический рефлекс. Рефлекс предупреждает разрывы мембран, снижая силу звука на 20дБ и более. Рефлекторное сокращение этих мышц наступает через 10мс после начала сильного звука, поэтому при мгновенных сильных раздражениях этот защитный механизм не успевает сработать, что может привести к нарушениям слуха. Стременная мышца кроме акустического рефлекса обеспечивает предвокализоционный рефлекс, который срабатывает всякий раз, когда человек начинает разговаривать. Мышца рефлекторно сокращается и, напрягая костное сочленение, демпфирует колебания слуховых косточек, отклоняет стремечко от овального окна, что приводит к снижению громкости собственного голоса. Если бы этого рефлекса не было, то человек «глох» бы от своей речи.
Функции внутреннего уха:
1) преобразование механических колебаний в нервные импульсы;
2) кодирование звука по силе и высоте.
Внутреннее ухо представлено улиткой и ограничено овальным и круглым окном. Вдоль 2,5 завитка улитки (длиной 35 мм) идут 3 лестницы, ограниченные двумя мембранами: верхняя (вестибулярная), средняя и нижняя (барабанная) (Рис. 13). Верхняя от средней отделяется эластичной мембраной, а средняя отделяется от нижней - жесткой или базальной мембраной. Все каналы улитки заполнены жидкостью. Верхний и нижний каналы улитки соединены геликотремой (на верхушке улитки) и заполнены перилимфой. Полость среднего канала не сообщается с другими двумя каналами и заполнена эндолимфой. Диаметр костного канала у основания улитки 0,04мм, а на вершине её - 0,5мм.

РиС. 13. Поперечный разрез завитка улитки (А) с увеличенной частью спирального (кортиев) органа (Б).
— секреторный эпителий; 2 — покровная мембрана; 3 — спиральный (кортиев) орган; 4 — спиральная связка; 5 - барабанная лестница; б - слой миелина; 7 — спиральный ганглий; 8 — капсула спирального ганглия; 9 — спиральная артерия; 10 —преддверная мембрана Рсйсснера;
—лестница преддверия: 12 — перепончатый канал; 13 - основная мембрана; 14 — внутренняя спиральная борозда; 15 — наружные волосковые клетки; 16 — решетчатая пластинка; 17 - нервные волокна, идущие к волосковым клеткам; 18 - внутренняя волосковая клетка.
МЕХАНИЗМ ПЕРЕДАЧИ ЗВУКОВЫХ КОЛЕБАНИЙ В СРЕДНЕМ И
ВНУТРЕННЕМ УХЕ.
Колебания барабанной перепонки передаются на слуховые косточки среднего уха: молоточек - наковальня - стремечко и дальше на овальное окно внутреннего уха..Колебания овального окна передаются перилимфе вестибулярной лестницы и через очень тонкую и поэтому не оказывающую сопротивления вестибулярную мембрану - эндолимфе (Рис.14), которая в свою очередь приводит в движение упругую базальную мембрану, а значит расположенный на ней - кортиев орган - рецепторный аппарат слуховой сенсорной системы.

Рис.14. Механизм передачи колебаний эндолимфы на покровную мембрану и рецепторные клетки кортиева органа
Кортиев орган представлен базальной мембраной, на которой расположены рецепторные клетки, имеющие волоски. Различают внутренние, расположенные в один ряд волосковые клетки (их 3500) и наружные, в 3-4 ряда (12-20 тыс.). Над волосками расположена покровная мембрана. Как только эндолимфа начинает колебаться, то покровная мембрана также перемещается сверху вниз, деформируя волоски рецепторных клеток. Волоски - стериоцилии (около 100 на каждом рецепторе), соединены в пучок поперечными нитями, так что наклон одной или нескольких стериоцилий приводит к сгибанию всех остальных. При этом чисто механически открываются К+ каналы на их мембранах и возникает РП. В дальнейшем К+ возвращается в сосудистую полоску, а дальше в эндолимфу. Деполяризация пресинаптического окончания волосковой клетки приводит к выходу в синаптическую щель нейромедиатора, который возбуждает постсинаптическую мембрану (веточка слухового нерва) и способствует появлению ГП и ПД. Сигналы от волосковых клеток по 32тыс. афферентных нервных волокнах - дендритах нейронов спирального ганглия поступают в составе слухового нерва в головной мозг. 90% волокон идёт от внутренних волосковых клеток и 10% - от наружных. Волна колебаний с ендолимфы переходит на перелимфу нижней лестницы. Из-за несжимаемости жидкости при колебаниях овального окна, выпячивается круглое окно.
Передача звука воздушным путём через наружное, среднее и внутреннее ухо, т.е. специализированную чувствительную к звуковым колебаниям систему, называется воздушной проводимостью. Кроме воздушного пути звуковая волна к кортиеву органу может поступать и через кости черепа. Убедиться в наличии костной проводимости можно достаточно легко, если поставить на темя ножку калебающегося камертона. Однако эффективность костного пути значительно ниже воздушного и имеет значение в случае восприятия очень громких звуков. Звук, передаваемый через кости, создаст волны сжатия и разрежения, вытесняя жидкость из вестибулярного лабиринта в улитку, а затем обратно. Кроме этого, колебания тимпапильно-косточкового аппарата в силу инерции отстают от колебаний, свойственных костям черепа. В результате стремя движется относительно каменистой кости, возбуждая внутреннее ухо- масооинерционная теория. При живой речи часть энергии передаётся в ухо по костям, поэтому в записи мы свой голос не узнаем.
ЗВУКОВЫЕ ПОРОГИ. ЗВУКОВАЯ ТРАВМА
Ухо человека воспринимает звук имеющий определенную силу, которая коррелирует с амплитудой звуковой волны. Амплитуду звуковой волны можно выразить как максимальное изменение давления на барабанную перепонку. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют - абсолютным порогом слуховой чувствительности. Ухо человека воспринимает диапазон от 0 до 140 дБ, который охватывает уровни от нижнего порога слухового ощущения, до такого, который может повредить кортиев орган, практически отображает десятимиллионную (107) разницу в давлении звуковых волн на барабанную перепонку, (Рис. 15).

Интенсивность самых распространенных звуков: (дБ)
160 реакционный самолет,боль
120 дискомфорт шум метро, живая рок-музыка
80 шум интенсивного уличного движения
нормальная речь
40 шепот
0 слуховой порог (0,0002 дин/см3)
Кроме силы звук различается по высоте. Высота звука коррелирует с частотой (количество колебаний за единицу времени). Человек воспринимает звуковые частоты от 20 до 20000 Гц (герц или колебаний за секунду). Слуховые пороги уха различаются для тонов разной высоты(рис.15).Для сравнения, тон мужского голоса равен приблизительно 120 Гц, тон женского голоса - 250 Гц. Значительно труднее различаются тоны высоких или низких частот. Так, порог слышимости на частоте 500Г'ц-20дБ, 200Гц-35дБ, 100Гц-60дБ. Такая малая чувствительность к звукам низкочастотного диапазона предохраняет человека от постоянных низкочастотных колебаний собственного тела (сосудов, сердца, мышц).
ТЕОРИИ РАЗЛИЧЕНИЯ ВЫСОТЫ И СИЛЫ ЗВУКА.
Максимальный уровень громкости (силы), вызывающий болевое ощущение равен 130-140дБ. Резкие, быстрые звуки такой силы могут привести к звуковой травме - разрыву барабанной перепонки. В таких случаях защитный акустический рефлекс, связанный с сокращением мышцы напрягающей барабанную перепонку не успевает сработать, т.к. он
наступает через 10мс после начала действия сильного звука. Кроме того, громкис звуки приводят к поражению волоскоиых клеток, их гибели и снижению слуха.
Предполагают наличие двух механизмов различия высоты тонов. Звуковая волна, представляющая из себя колебание молекул воздуха, распространяется в виде продольной волны давления. Передаваясь на пери- и эндолимфу, эта бегущая волна между местом возникновения и затухания имеет участок, где амплитуда колебаний максимальна (явление резонанса) место нахождения этого амплитудного максимума зависит от частоты колебания: при более высоких частотах он ближе к овальной мембране, а при низких - ближе к геликотреме (Рис.16) (место перехода верхней лестницы в нижнюю).
Рис. 16. Теория бегущей волны.

В результате резонансный амплитудный максимум для каждой частоты располагается в специфической точке эидолимфатичсского капала.Расположенные в этом месге волокна базилярной мембраны и лежащие на них волосковые клетки колеблются наиболее сильно, так что по нервным волокнам, отходящих от них, идет наиболее мощная импульсация. Это пр остранственпая теория кодирования высоты тона в самом рецепторе - теория «бегущей волны», или теория «места». Бегущая волна имеет наибольшую амплитуду на строго определённом участке мембраны в засимосги ог частоты. Градиент жёсткости мембраны, который увеличивается от овального окна к гсликотреме, всегда заставляет волны двигаться в том же направлении и никогда в обратном.
При небольшой частоте колебаний (до 1кГц) может использоваться и телефон ный механизм кодирования, который предполагает, что частота колебаний потенциалов в слуховом нерве при восприятии звуков разной высоты соответствует частотам воспринимаемых звуков, как это происходит в телефонном кабеле при передаче звуков по телефону. Так как рефракторный период быстро проводящих волокон не бывает ниже I сек, следовательно, информация о частоте звука в виде синхронных со /ком колебаний потенциала, т.е. телефонный принцип, может распрострапяться лишь на звуковые частоты до 1000Гц. Т.о. при действии низких и средних тонов кроме пространственного осуществляется и временное кодирование. Для каждого нейрона существует оптимальная, и характеристическая частота звука, на которую порог реакции нейрона минимален, а в обе стороны по диапазону частот от этого оптимума порог резко возрастает. Т.о. каждый нейрон настроен на выделение из всей совокупности звуков лишь определенного, достаточно узкого участка частотного диапазона.
В рецепторах волосковых клеток звук кодируется по силе. Различение силы звука зависит от количества возбужденных рецепторных клеток кортиева органа. Чем больше сила звука, тем больше амплитуда колебаний эндолимфы, тем больше амплитуда колебаний мембран средней лестницы. А следовательно, большее число волосковых клеток органа Корти вовлекается в возбуждение. Кроме того, пороги возбуждения внутренних и наружных волосковых клеток разные: НП внутренних возникают при большей силе звука. В различении звука по силе имеет место и частота импульсации - т. е. чем чаще разряжаются волосковые клетки, тем сильнее звук.
ЭНДОКОХЛЕАРНЫЙ И МИКРОФОННЫЙ ПОТЕНЦИАЛ УЛИТКИ
При отведении электрических потенциалов от улитки было обнаружено несколько электрических феноменов. Два из них - мембранный потенциал слуховой рецепторной клетки и потенциал эндолимфы не связаны с действием звука, т.е. наблюдаются и при его отсутствии. А микрофонный потенциал улитки возникает иод влиянием звуковых раздражений.
Между эндолимфой и внутренней частью мембраны слуховой рецепторной клетки формируется МП равный 150 мВ. Такой МП обеспечивает высокую чувствительность реценторных клеток к слабым звуковым колебаниям и называется эндохлеарным. Эндолимфа, которую продуцирует сосудистая полоска, содержит в 100 раз больше К' ив 10 раз меньше Nа', чем в перелимфе. Это обеспечивается высоким уровнем Nа', К + - А'ТФазы, а так же К'-помпы. Поэтому эндолимфа заряжена положительно по отношению к перелимфе, где разность потенциалов

она больше 3000Гц, то решающую роль в определении источника звука не играет разница в силе звука. Так, при высоких частотах, когда величина головы значительно больше длины звуковой волны, последняя не может «огибать» это препятствие, звук отражается головой. В результате возникает разница в интенсивности звуков, поступающих на правое и левое ухо. У человека дифференциальный порог но интенсивности на частоте 1000Гц равен IдБ, т.е. очень высокий. Поэтому различение интенсивности будет тонкое, что и позволит определить источник звука.
ОБРАБОТКА ЗВУКОВОЙ ИНФОРМАЦИИ В ПОДКОРКОВЫХ И КОРКОВЫХ ЦЕНТРАХ
Нейроны первого порядка слухового пути входят в состав спирального ганглия улитки, центральные отростки которого образуют кохлеарный (слуховой) нерв. Слуховой нерв, являясь ветвью VIII пары черепно- мозговых нервов, проходит в продолговатый мозг и заканчивается на кохлеарных ядрах (второй нейрон) (Рис 17). Затем слуховой путь делится на ипси- и, более мощный, контрлатеральный пучок волокон, который достигает олив Аксоны олив ипси- и контрлатеральных волокон проходят в составе латеральной петли Одна часть петли проходит в нижние буг ры четверохолмия, другая - не переключаясь, в медиальное коленчатое тело тялпмуся. Конечные ядра находятся в первичной и вторичной слуховой зоне височной доли мозга (41 и 42 поле по Бромапу).
Информация, содержащаяся в звуковом сигнале, пройдя все указанные ядра, многократно «переписывается». При этом, на каждом этапе происходит ее соответствующий анализ, причем, нередко с подключением других «не слуховых» отделов ЦНС.
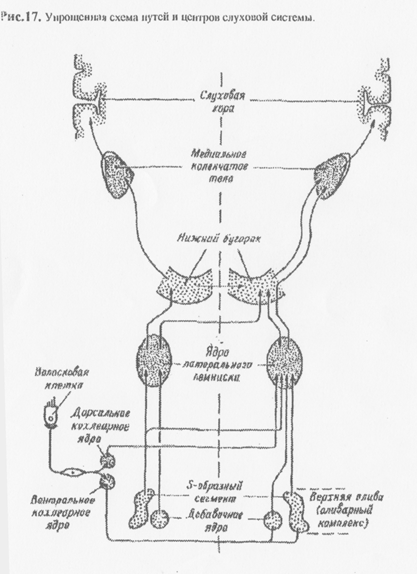
Нейроны вентрального ядра еще воспринимают чистые тоны, т.е.возбуждение в них возникает при действии строго определенных частот. В дорсальном ядре, кроме реакции на простой стимул, возникают возбуждения на более сложные переменные частоты, прекращение звука ит.п. На более высоких уровнях в нейронах постепенно увеличивается специфичность реагирования на сложные звуковые модуляции. Таким образом, каждый раз в нервных слуховых центрах возникает своеобразная мозаика возбуждающихся нейронов (одни возбуждаются при меняющейся частоте, другие - меняющейся амплитуде, третьи - при варьировании длительности расстояния от источника, его перемещении и т.п.) Наиболее полно и совершенно эти процессы происходят лишь в корковых огделах. Корковые нейроны, воспринимающие низкие тоны, находятся в иереднелатеральной части слуховой зоны, а высокие - в заднемедиальной. Одни нейроны активируются противоположным ухом, другие - от
одноименного уха, третьи - только при одновременной стимуляции от обоих ушей. У людей, особенно правшей, больше развита левая часть височной закрутки, что связано с рече-слуховым восприятием. Интересно, что у музыкантов этот участок превышает свои размеры.
Дата добавления: 2015-08-17; просмотров: 177 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| ПОЛЕ ЗРЕНИЯ | | | Российские нормативно-правовые акты деятельности по оказанию гостиничных услуг |