Читайте также:
|
Химическая классификация
1. Сложные эфиры – ацетилхолин (АХ).
2. Биогенные амины: 1) катехоламины (дофамин, норадреналин (НА), адреналин (А)); 2) серотонин; 3) гистамин.
3. Аминокислоты: 1) гаммааминомасляная кислота (ГАМК); 2) глутаминовая кислота; 3) глицин; 4) аргинин.
4. Пептиды: 1) опиоидные пептиды (а)метэнкефалин; б) энкефалины; в) лейэнкефалины; 2) вещество «Р»; 3) вазоактивный интестинальный пептид; 4) соматостатин.
5. Пуриновые соединения: АТФ.
6. Вещества с минимальной молекулярной массой: 1) N0; 2) СО.
Функциональная классификация
1. Возбуждающие медиаторы, вызывающие деполяризацию постсинаптической мембраны и образование возбуждающего постсинаптического потенциала: 1) АХ; 2) глутаминовая кислота; 3) аспарагиновая кислота.
2. Тормозящие медиаторы, вызывающие гиперполяризацию постсинаптической мембраны, после чего возникает тормозной постсинаптический потенциал, который генерирует процесс торможения: 1) ГАМК; 2) глицин; 3) вещество «Р»; 4) дофамин; 5) серотонин; 6) АТФ.
Норадреналин, изонорадреналин, адреналин, гистамин являются как тормозными, так и возбуждающими.
АХ (ацетилхолин) является самым распространенным медиатором в ЦНС и в периферической нервной системе. Содержание АХ в различных структурах нервной системы неодинаково. С филогенетической точки зрения в более древних структурах нервной системы концентрация ацетилхолина выше, чем в молодых. АХ находится в тканях в двух состояниях: связан с белками или находится в свободном состоянии (активный медиатор находится только в этом состоянии).
АХ образуется из аминокислоты холин и ацетил-коэнзима А.
Медиаторами в адренэргических синапсах являются норадреналин, изонорадреналин, адреналин. Образование катехоламинов идет в везикулах терминали аксона, источником является аминокислота: фенилаланин (ФА).
Возбуждающие постсинаптические потенциалы (ВПСП). Тормозные постсинаптические потенциалы (ТПСП). Миниатюрные потенциалы.
Медиатор, находящийся в пузырьках, выделяется в синаптическую щель с помощью экзоцитоза, (пузырьки подходят к мембране, сливаются с ней и разрываются, выпуская медиатор). Его выделение происходит небольшими порциями - квантами. Каждый квант содержит от 1.000 до 10.000 молекул неиромедиатора. Небольшое количество квантов находит из окончания и в состоянии покоя. Когда нервный импульс, т.е. ПД, достигает пресинаптического окончания, происходит деполяризация его пресинаптической мембраны. Открываются ее кальциевые каналы и ионы кальция «ходят в синаптическую бляшку. Начинается выделение большого количества квантов неиромедиатора. Молекулы медиатора диффундируют через синаптическую щель к постсинаптической мембране и взаимодействуют с её хеморецепторами. В результате образования комплексов медиатор-рецептор, в субсинаптической мембране начинается синтез так называемых вторичных посредников, В частности цАМД). Эти посредники активируют ионные каналы постсинаптической мембраны. Поэтому такие каналы называют хемозависимыми или рецепторуправляемыми, т.е они открываются при действии-ФАВ на хеморецепторы. В результате открывания каналов изменяется потенциал субсинаптической мембраны. Такое изменение называется постсинаптическим потенциалом.
В ЦНС возбуждающими являются холин, адрен, дофамин, серотонинергические синапсы, и некоторые другие. При взаимодействий их медиаторов с соответствующими рецепторами, открываются хемозависимые натриевые каналы. Ионы натрия входят в клетку через субсинаптическую мембрану. Происходит ее местная или распространяющаяся деполяризация. Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП). Тормозными являются глицин- и ГАМКергические синапсы. При связывании медиатора с хеморецепторами. активируются калиевые или хлорные хемозависимые каналы. В результате ионы калия выходят из клетки через, мембрану. Ионы хлора входят через нее. Возникает только местная гиперполяризация субсинаптической мембраны. Она называется тормозным постсинаптическим потенциалом (ТПСП).
Величина ВПСП и ТПСП определяется количеством квантов медиатора, выделившихся из терминали а, следовательно, частотой нервных импульсов, т.е синоптическая передача не подчиняется закону "все или ничего".
Если количество выделившегося возбуждающего медиатора достаточно велико, то в субсинаптической мембране может генерироваться распространяющийся ПД. ТПСП, независимо от количества медиатора не распространяется за пределы субсинаптической мембраны.
После прекращения поступления нервных импульсов, выделившийся медиатор удаляется из синаптической щели тремя путями:
1. Разрушается специальными ферментами, фиксированными на поверхности субсинаптнческой мембраны. В холинэргических синапсах это ацетилхолннэстераза (А-ХЭ). В адренергических, дофаминергических, серотонинергических моноаминоксидаза (МАО) и катехол-о-метилтрансфераза (КОМТ).
2. Часть медиатора возвращается в пресинаптическое окончание с помощью процесса обратного захвата (значение в том что синтез нового нейромедиатора длительный процесс).
3. Небольшое количество уносится межклеточной жидкостью.
Особенности передачи возбуждения через химические синапсы:
1. Возбуждение передается только в одном направлении, это способствует его точному распространению в ЦНС.
2. Они обладают синаптической задержкой. Это время необходимое на выделения медиатора, его диффузию и процессы в субсинаптической мембране.
3. В синапсах происходит трансформация, т.е. изменение частоты нервных импульсов.
4. Для них характерно явление суммации. Т.е. чем больше частота импульсов, тем выше амплитуда ВПСП и ТПСП.
5. Синапсы обладают низкой лабильностью.
Периферические синапсы образованы терминалями эфферентных нервов и участками мембран исполнительных органов. Например, нервно-мышечные синапсы образуются окончаниями аксонов двигательных нейронов и мышечными волокнами. Благодаря своеобразной форме они называются нервно-мышечными концевыми пластинками. Их общий план строения такой же, как у всех химических синапсов, но субсинаптическая мембрана толще и образует многочисленные субсинаптические складки. Они увеличивают площадь синаптического контакта. Медиатором этих синапсов является ацетилхолин.
В субсинаптическую мембрану встроены Н-холинорецепторы, т.е. холинорецепторы, которые помимо АХ могут связываться и с никотином. Взаимодействие холинорецепторами приводит к открыванию хемозависимых натриевых каналов и развитию деполяризации. В связи с тем, что отдельные кванты ацетилхолина выделяется и в состоянии покоя, в постсинаптической мембране нервно-мышечных синапсов постоянно возникают слабые кратковременные всплески деполяризации - миниатюрные потенциалы концевой пластинки (МПКП) При поступлении нервного импульса, выделяется большое количество АХ и развивается выраженная деполяризация, называемая потенциалом концевой пластинки (ПКП). В отличие от центральных, в нервно-мышечных синапсах ПКП всегда значительно выше критического уровня деполяризации. Поэтому он всегда сопровождается генерацией ПД и сокращением мышечного волокна. Т.е. для распространяющегося возбуждения и сокращения суммации эффектов квантов нейромедиатора не требуется.
Ядкураре и курареподобные препараты фармакологические препараты резко снижают ПКП и блокируют нервно-мышечную передачу. В результате выключается вся скелетная мускулатура, в том числе и дыхательная. Это используется для операций с искусственной вентиляцией легких. Разрушение АХ осуществляется ферментом ацетилхолинестеразой. Некоторые фосфороорганические вещества (хлорофос, зарин) инактнвируют холинэстеразу. Поэтому АХ накапливается в синапсах и возникают мышечные судороги. В клинической практике известно заболевание myasthenia gravis при котором уменьшается количество колинорецепторов в концевых пластинках. Это связано с выработкой аутоиммунных антител к ним. В результате амплитуда ПКП значительно снижается. Возникают резкая слабость и патологическая утомляемость.
Современные представления о передаче возбуждения в синапсах.
Синапс (греч. synapsis соприкосновение, соединение) — специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически синапс образован контактирующими мембранами двух клеток. Мембрана, принадлежащая отросткам нервных клеток, называется пресинаптической, мембрана клетки, к которой передается сигнал, — постсинаптической.
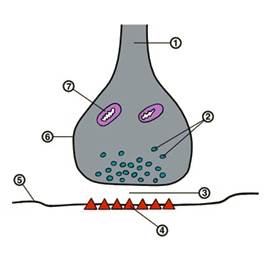
Схема строения межнейронного синапса: 1 — нервное волокно (аксон); 2 — везикулы или синаптические пузырьки; 3 — синаптическая щель; 4 — рецепторы для медиатора; 5 — постсинаптическая мембрана; 6 —пресинаптическая мембрана; 7 — митохондрия.
В соответствии с принадлежностью постсинаптической мембраны синапсы подразделяют на нейросекреторные, нейромышечные и межнейроннные. Последние в зависимости от места их расположения разделяют на аксодендритические, аксосоматические, аксо-аксональные и дендро-дендритические. Различают аксодендритные синапсы (синапс между концевыми веточками аксона одного нейрона и дендритами другого нейрона) и аксосоматические синапсы (синапс между концевыми веточками аксона одного нейрона и телом другого нейрона).
В зависимости от природы проходящих через синапсы сигналов, синапсы делятся на электрические синапсы (так называемыеэфапсы) и химические синапсы. В синапсах с химической передачей возбуждения между пре- и постсинаптической мембранами имеется синаптическая щель, куда выделяется химическое вещество-передатчик — медиатор. Химические синапсы часто обозначают по названию медиатора (например, холинергические, адренергические, серотонинергические и т.п.). В эфапсе пре- и постсинаптические мембраны плотно соприкасаются и возбуждение передается посредством электрического тока. В зависимости от изменения биопотенциала постсинаптической мембраны различают синапсы деполяризующие, или возбуждающие, и гиперполяризующие, или тормозные.
 Число синапсов очень велико, что обеспечивает большую площадь для передачи информации. На дендритах и телах отдельных двигательных нейронов спинного мозга находится свыше 1 000 синапсов. Некоторые клетки головного мозга могут иметь до 10 000 синапсов.
Число синапсов очень велико, что обеспечивает большую площадь для передачи информации. На дендритах и телах отдельных двигательных нейронов спинного мозга находится свыше 1 000 синапсов. Некоторые клетки головного мозга могут иметь до 10 000 синапсов.
Механизм передачи возбуждения
Механизм передачи возбуждения принципиально одинаков во всех химических синапсах. В нем можно выделить следующие основные этапы: синтез и депонирование медиатора в пресинаптическом нейроне и его окончаниях; высвобождение медиатора из депонирующих везикул и его выход в синаптическую щель; взаимодействие медиатора со специфическими хеморецепторами постсинаптической мембраны с последующей генерацией биоэлектрического потенциала; инактивация выделенного медиатора с помощью ферментов или системы обратного поглощения. Биологически активные вещества, выполняющие функцию медиаторов, делят на несколько групп.
К классическим нейромедиаторам относят ацетилхолин, адреналин и норадреналин, дофамин, серотонин, а также аминокислоты глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды. В соответствии с принципом Дейла, каждый отдельный нейрон в своих синаптических окончаниях один и тот же медиатор, поэтому нейроны можно обозначать также по виду медиатора: холинергические, адренергические, ГАМК-эргические, пептидергические. Синтез нейромедиаторов осуществляется как в соме нейрона с последующим аксонным транспортом, так и непосредственно в пресинаптических окончаниях аксона, где медиатор концентрируется в везикулах, или синаптических пузырьках.
В состоянии функционального покоя в пресинаптическом окончании происходит случайный контакт синаптических пузырьков с пресинаптической мембраной и выделение в синаптическую щель порции (кванта) медиатора из отдельной везикулы. Выделившийся в синаптическую щель медиатор взаимодействует с хеморецепторами постсинаптической мембраны и приводит к возникновению миниатюрного постсинаптического потенциала. Приходящий к пресинаптическому окончанию потенциал действия в несколько раз увеличивает количество выделяемого в синаптическую щель медиатора. Взаимосвязь между потенциалом действия пресинаптической мембраны и процессом выделения медиатора из везикул в синаптическую щель обеспечивается ионами Са++. Выделяющийся в синаптическую щель медиатор взаимодействует с различными хеморецептивными участками на постсинаптической мембране. Роль мембранных, или клеточных, рецепторов играют белковые молекулы, обладающие способностью «узнавать» специфические для них вещества и вступать с ними в реакцию. В этом процессе большое значение придается системе аденилатциклаза-циклический АМФ.
Предполагается, что взаимодействие медиатора с рецепторным участком аденилатциклазы приводит к образованию циклического АМФ с последующим повышением активности протеинкиназ цитоплазмы и ядра клетки. Вследствие этого интенсифицируется фосфорилирование белков, белковые молекулы подвергаются конформационным изменениям и происходит активация специальных ионных каналов мембраны. При увеличении проницаемости постсинаптической мембраны для ионов натрия, калия и хлора возникает ее деполяризация, регистрируется возбуждающий постсинаптический потенциал (ВПСП). При увеличении проницаемости лишь для ионов калия и хлора мембрана гиперполяризуется и регистрируется тормозный постсинаптический потенциал (ТПСП). ВПСП могут суммироваться, и при достижении величины деполяризации мембраны критического уровня генерируется потенциал действия. ТПСП тормозит генерацию потенциала действия, уменьшая суммарную величину ВПСП.
Синапс, его виды. Понятие о медиаторах. Опыт Леви.
Дата добавления: 2015-08-02; просмотров: 67 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Промывание желудка с использованием толстого зонда | | | Устройство и работа синапса |