Читайте также:
|
Клиницистам хорошо известен тот факт, что больные с массивными поражениями лобных долей мозга находятся, как правило, в состоянии сниженной активности, что их внимание легко отвлекается любым побочным раздражителем и что организовать их внимание, прочно подчинив его вызванному намерению, обычно невозможно.
Этот основной факт много раз описывался невропатологами и психиатрами, наблюдавшими больных с массивными поражениями лобных долей мозга (Клейст, 1908, 1930, 1934; Фейхтвангер, 1923; Райлендер, 1939; Леонхардт, 1964, 1965, Экаэн и Ажуриагерра, 1956), но не был, однако, предметом тщательного систематического исследования.
Лишь за последние годы — в связи с успехами нейрофизиологии, указавшими на теснейшие взаимоотношения между лобными долями мозга и стволовой и таламической ретикулярной формацией (ом. выше II, 5, б), а также в связи с введением в клинику точных нейропсихологических и психофизиологических исследований, научный анализ той роли, которую играют лобные доли мозга в регуляции состояний активности, стал более доступен.
Значительная часть этих исследований, проведенных Е. Д. Хомской и ее сотрудниками, была посвящена изучению ориентировочных реакций и анализу той роли, которую лобные доли мозга играют в их регуляции.
Данные, полученные при этих исследованиях, подвергнуты детальному анализу в другом месте (А Р. Лурия и Е. Д. Хомская, 1966), и здесь мы остановимся на их изложении лишь в самом общем виде.
Известно, что каждый новый или значимый раздражитель вызывает ориентировочный рефлекс или реакцию активации (arousal). Эта реакция активации приводит к усилению бодрственного состояния сознания и выражается в целом ряде изменений дыхания, кровенаполнения сосудов, кожно-гальванической реакции (которые относятся к числу вегетативных компонентов ориентировочного рефлекса), а также в хорошо известных в электрофизиологии явлениях депрессии альфа-ритма и в ряде специальных форм изменения электрической активности, на которых оды еще остановимся ниже.
Эти симптомы ориентировочного рефлекса (или признаки активации) сохраняются некоторое время, однако при повторении тех же раздражителей постепенно угасают. Такое угасание различных компонентов ориентировочного рефлекса (или явление привыкания — habituation — англо-американских авторов) наступает уже после 10— 15-кратного повторения одного и того же раздражителя и устраняется лишь при предъявлении нового раздражителя или изменении каких-либо параметров старого раздражителя.
Существенное для нормального человека явление, детально изученное Ε. Η. Соколовым (1957, 1958, 1959, 1960 и др.), О. С. Виноградовой (1959 и др.), заключается в том, что восстановление вегетативных компонентов угасшего ориентировочного рефлекса можно вызвать и иным путем. Для этого достаточно посредством речевой инструкции придать данному раздражителю сигнальное значение, например, предложить испытуемому считать предъявляемые сигналы, следить, не наступят ли в них какие-либо изменения, или отвечать на каждый сигнал какой-либо двигательной или словесной реакцией. В этих случаях, как это было обнаружено в исследованиях E. Д. Хамской (1960, 1961, 1963, 1965), вегетативные компоненты ориентировочного рефлекса восстанавливаются и длительное время перестают проявлять тенденцию к угасанию; таким образом, у испытуемого возникает стойкое состояние активации. Симптомы такой длительной активации коры с помощью речевой инструкции можно видеть у нормального испытуемого в опытах с записью плетизмографических и кожно-гальванических реакций на раздражители, которым с помощью инструкции придается сигнальное значение. На рис. 71 мы приводим примеры, взятые из соответствующих опытов Е. Д. Хамской.

Рис. 71. Восстановление кожно-гальваниче-ского компонента ориентировочного рефлекса на звуки с помощью речевой инструкции у нормальных испытуемых (по Е. Д. Хомской): а — угашение кожно-гальванической реакции на прерывистые звуки (60 дб) у трех испытуемых. На рисунках представлены три типа угашения: А — быстрое угашение, Б — длительное угашение, В — отсутствие КГР на звуки. Цифры обозначают порядковые номера раздражителей; б — восстановление ориентировочных кожно-гальванических реакций после введения инструкции считать количество звуков в каждом прерывистом раздражителе. Стрелка обозначает введение инструкции. «PO»—речевой ответ испытуемого; в — исчезновение ориентировочных кожно-гальванических реакций после отмены задания
Восстановление ориентировочной реакции на раздражитель, которому придано сигнальное значение, может стать настолько устойчивым и специфическим, что у нормального испытуемого ориентировочные реакции на все побочные раздражители могут затормозиться, и даже физически более сильные раздражители, не имеющие сигнального значения, могут перестать вызывать сколько-нибудь отчетливые вегетативные компоненты ориентировочного рефлекса. На рис. 72 мы приводим такой пример, взятый из опытов О. С. Виноградовой.

Р ис. 72. Торможение сосудистых реакций на побочные раздражители после формирования доминантного очага с помощью речевой инструкции (по О. С Виноградовой) А. Опыт с нормальным испытуемым. Слабые сигнальные звуки вызывают стойкую ориентировочную реакцию (верхняя строка); сильный стук за окном не вызывает реакции (нижняя строка). Б и В. Опыты с олигофренами. Слабые сигнальные звуки быстро перестают вызывать реакцию (обе верхние строки). Сильный стук за окном вызывает выраженную реакцию (обе нижние строки)
Характерные особенности поведения вегетативных компонентов ориентировочного рефлекса при патологических состояниях мозга были описаны Е. Д. Хамской (1960, 1961, 1963, 1966), Н. А. Филиппычевой (1960, 1966) и М. Климковским (1961).
Как показали эти исследования, у больных с очаговыми поражениями (опухолями) мозга могут проявляться заметные патологические изменения вегетативных компонентов ориентировочного рефлекса. У одних таких больных вегетативные компоненты ориентировочного рефлекса могут быть слабо выражены, а иногда и совсем угнетены, у других они могут быть патологически усилены и иногда не угасать в течение длительного срока. Характерная черта заключается и том, что если речевая инструкция, придающая раздражителям сигнальное значение, приводит у больных с поражением задних (теменно-затылочных и височных) отделов мозга к стойкому восстановлению ориентировочного рефлекса, то у больных с поражением лобных отделов мозга такое влияние речевой инструкции патологически изменено: плохо выраженные вегетативные компоненты ориентировочного рефлекса либо вовсе не восстанавливаются после того, как речевая инструкция придает раздражителям сигнальное значение, либо же (в случаях менее выраженного «лобного синдрома») это восстановление вегетативных компонентов ориентировочного рефлекса носит диффузный и нестойкий характер.
Иллюстрации этого факта приводятся на рисунках 73 и 74, взятых из исследований Е. Д. Хомской.

Рис. 73 Восстановление вегетативных компонентов ориентировочного рефлекса на звуки с помощью речевой инструкции у больного с поражением правой теменной области больших полушарий (по E. Д. Хомской) α — угашение вегетативных компонентов ориентировочного рефлекса на прерывистые звуки (70 дб); верхняя кривая — сосудистые реакции; нижняя кривая — кожно-гальванические реакции Цифры обозначают интенсивность звуков (в дб) и их количество в пачке; б — восстановление вегетативных компонентов ориентировочного рефлекса после введения инструкции считать количество звуков.

Рис. 74. Различные типы нарушения активирующего влияния инструкции («считать звуки») на сосудистые и кожно-гальванические ориентировочные реакции у больных с массивным поражением лобных долей мозга: /—// — отсутствие каких бы то ни было изменений в плетизмограмме и КХР при предъявлении индифферентных звуков (/) и при их сосчитывании (//). Нет изменений тонической формы КГР; больной 3. (парасагиттальная арахноидэндотелиома левой лобной доли, глубоких ее отделов, прилегающая к серповидному отростку и вторично воздействующая на мозолистое тело); ///—IV—влияние инструкции «считать звуки» только на тоническую форму КГР; ориентировочные сосудистые реакции и КГР как на индифферентные (///), так и на сигнальные (/V) звуки отсутствуют; больной Н. (очень больших размеров арахноидэндотелиома малого крыла основной кости справа—7X7X8 см); V—VI—появление в фоне КГР после введения инструкции (VI) не только тонических изменений, но и частых спонтанных колебаний, которых не было до инструкции (V). Отчетливого сужения сосудов и изменения кожного потенциала при сосчитывании звуков в пачке нет; больная А. (дедиф-ференцирующая астроцитома полюса левой лобной доли с кистой).
Они показывают, что лобные доли мозга играют существенную роль в сложных формах регуляции ориентировочного рефлекса и позволяют ближе подойти к механизмам того нарушения состояний активации, которые описывали клиницисты, наблюдая больных с массивными поражениями лобных долей мозга. Тот факт, что речевая система перестает в этих случаях участвовать в формировании прочных и избирательных состояний активности, которые лежат в основе любых сложных форм целенаправленного (поведения, раскрывает важную сторону патологии лобных долей мозга и позволяет сделать важные выводы об их функциональном значении.
Факты, указывающие на то, что лобные доли мозга играют важную роль в возникновении и сохранении активных состояний мозговой коры и связанных с ними форм активного поведения, выступают и при ряде электрофизиологических исследований.
Как показали наблюдения Грей Уолтера (1966), всякое ожидание сигнала вызывает в коре головного мозга человека своеобразные медленные волны, которые автор назвал «волнами ожидания» (expectancy waves). Эти медленные изменения потенциалов, возникающие в ответ на инструкцию ожидать появления условного раздражителя, являющегося сигналом для движения, Грей Уолтер наблюдал исключительно в лобных долях мозга; интенсивность таких «волн ожидания» менялась в зависимости от степени вероятности появления сигнала; при отмене инструкции ожидать сигнал эти волны исчезали (рис. 75).
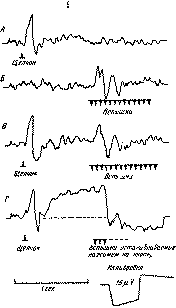


Рис. 75 Динамика «волн ожидания» в раз личных условиях опыта (по Грей Уолтеру, 1966)см. на стр. 229 230, 231) 1 — Сравнение ответов А — изолированные звуковые сигналы, Б — изолированные зри тельные сигналы, В — ассоциация сигналов, Г — сочетание сигьалов с реакцией испытуемого «Волны ожидания» наблюдаются лишь в последнем случае II — Влияние устранения пускового сигнала А — контрольная связь с «волнами ожидания», Б — усредненная запись первых шести предъявлений без пускового сигнала, «волны ожидания» редуцируются, видна условная реакция в тот момент, когда ожи дается появление пускового сигнала, В, Г, Д — последовательная усредненная реги страция, показывающая постепенное исчезновение «волн ожидания», E — быстрое воcстановление «волн ожидания» при повторном предъявлении пускового сигнала От метим, что данный испытуемый может быть отнесен к «сангвиникам», так как он легко приспосабливается к изменчивой ситуации III—Влияние установки (подготовки к сигналу) на появление «волн ожидания» А — испытуемый решил не нажимать на кнопку, «волны ожидания» исчезают, Б — испытуемый предупреждается, что пусковых сигна лов не будет «волны ожидания» сразу же исчезают, без инструкции они угасают через 30—50 предъявлений (см II), В — ис пытуемый ложно предупреждается о том, что вспышки появятся снова «Волны ожи дания» указывают на компромисс между доверием к инструкции и непосредственные опытом.
Эти факты (вместе с уже цитированными выше фактами, полечен ными К Прибрамом при изучении «реакции ожидания» у обезьян и ее изменении при экстирпации лобных долей) еще раз указывают на ту роль, которую играют лобные доли мозга в обеспечении устойчивых активных состояний организма
Близкие к этому факты были получены M H Ливановым и его сотрудниками (M H Ливанов, H А Гаврилава и А С Асланов, 1964, 1965, 1966) Как показали эти исследователи, регистрировавшие одновременно электрическую активность большого числа точек мозговой коры, всякая активная деятельность человека (например, решение сложных арифметических примеров) вызывает значительное увеличение числа синхронно работающих пунктов коры головного мозга, причем заметное повышение корреляции биоэлектрической активности работающих пунктов появляется прежде всего в лобных долях мозга
Аналогичные данные можно наблюдать и у больных с параноидной формой шизофрении, у которых непрерывное состояние напряжения, связанное с застойными пунктами возбуждения, проявляется также в повышении числа синхронно работающих пунктов лобной коры Было отмечено также, что применение аминазина, снижающего это напряженное состояние, приводит к тому, что только что отмеченное высокое число синхронно работающих пунктов лобной коры резко уменьшается.
Рис. 76 (стр. 232) иллюстрирует эти положения.

Рис. 76. Корреляция биопотенциалов работающих пунктов лобной коры при различных состояниях напряжения (по М. Н. Ливанову, Н. А. Гавриловой и А. С. Асланову, 1966) Вверху — Временная корреляция биопотенциалов точек коры головного мозга здорового испытуемого С. Е.: а —в состоянии покоя; б —на пятой секунде решения арифметической задачи. Кружками на карте коры головного мозга обозначены электроды, наложенные на поверхность головы. Полностью заштрихованы электроды, расположенные в передне-лобных отделах; сеткой — в области двигательного анализатора; вертикальными штрихами — в задних отделах коры. Далее на всех рисунках обозначения те же. Стрелками соединены участки коры, коррелирующие в высоком проценте времени Внизу — Временная корреляция биоэлектрической активности точек коры головного мозга больного параноидной формой шизофрении при применении аминазина; α — во время решения арифметической задачи до применения аминазина; б — во время решения аналогичной задачи через 1 час 15 мин после приема аминазина.
Легко видеть, что и эти факты показывают, что лобные доли мозга, имеющие, как было указано выше, интимную связь с ретикулярной формацией, играют существенную роль в обеспечении активных состояний организма, являясь важной составной частью наиболее сложных механизмов активации.
Все эти факты заставляют с особенным (вниманием отнестись к тем наблюдениям, которые указывают на снижение или патологическое изменение электрофизиологичеcких признаков наиболее сложных форм активации, наступающих гари массивных поражениях лобных долей мозга.
Такие факты были получены в последние годы Е. Д. Хомской и ее сотрудниками.
Как хорошо известно, всякое раздражение, доходящее до субъекта, вызывает изменения электроэнцефалограммы, принимающие обычно характер депрессии альфа-ритма. Эти явления, которые с полным основанием могут быть расценены как один из компонентов ориентировочного рефлекса, отчетливо наблюдаются в норме и выступают особенно резко, если действующий раздражитель оказывается достаточно неожиданным или если он приобретает сигнальное значение. Аналогичные факты проявляются и три исследовании тех больных с очаговыми поражениями задних отделов мозта, у которых альфа-ритм остается достаточно сохранным.
Иная картина, как это показали опыты О. П. Барановской и Е. Д. Хомской (1966), обнаруживается у больных с поражениями лобных отделов мозга (см. рис. 77). Как было установлено в этих наблюдениях, если у нормальных субъектов речевая инструкция, придающая сигнальное значение раздражителям, вызывает заметное усиление депрессии прежде «сего высокочастотных колебаний, входящих в группу альфа-частот (11—13 гц), и делает эту депрессию более длительной, то у больных с поражением лобных долей мозта введение речевой инструкции не приводит к таким изменениям электрической активности; у больных этой группы реакция активации либо полностью выпадает, либо они продолжают реагировать на сигнальные раздражители изменениями альфа-ритма той же интенсивности и длительности, какая отмечалась и на индифферентные 'раздражители, причем эти изменения носят нестойкий характер и проявляются главным образом в диапазоне более низких альфа-частот (8—9 гц).
Подобное действие речевой инструкции, вводящей сигнальное значение раздражителей, еще раз указывает на участие лобных долей в регуляции состояний активности.
Рис. 77 (стр. 233—235) иллюстрирует эти факты.

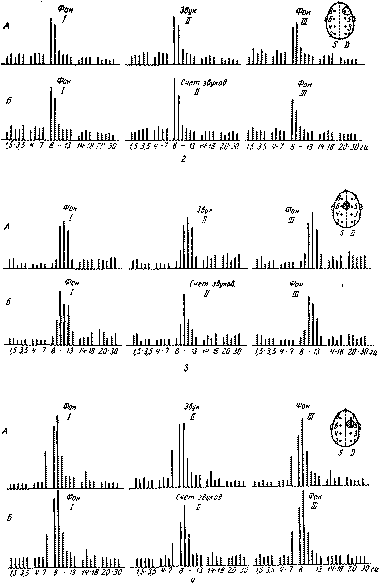

Рис. 77. Изменение частотного спектра электроэнцефалограммы под влиянием инструкции в норме и при поражении лобных долей мозга (по О. П. Барановской и Е. Д. Хомской, 1966): а — изменения частного спектра ЭЭГ при действии индифферентных и сигнальных звуковых раздражителей у здоровой испытуемой К. А — частотные спектры ЭЭГ перед звуками (/), в момент предъявления индифферентных звуков (II) и после их прекращения (///); Б — частотные спектры ЭЭГ перед звуками (/), в момент предъявления сигнальных звуков (//) и после их прекращения (///); В — частотные спектры ЭЭГ перед звуками (/), во время действия вновь индифферентных звуков (//) и после их прекращения {III). Каждый спектр — среднее из 15 периодов (анализатор Уолтера); б— изменения частотного спектра ЭЭГ, записанной с затылочно-теменных отделов мозга здорового полушария при предъявлении индифферентных и сигнальных раздражителей у больных с поражением лобных долей мозга. 1 — частотные спектры ЭЭГ больной Ф. (арахноидэндотелиома межполушарной щели в задних отделах лобных долей мозга, больше слева); 2 — частотные спектры ЭЭГ больного Д. (саркомотозная арахноидэндотелиома правой лобной кости, воздействующая на мозговое вещество лобной доли); 3 — частотные спектры ЭЭГ больного П. (двусторонняя парасагитталь-ная арахноидэндотелиома задних отделов лобных долей мозга); 4 — частотные спектры ЭЭГ больного Ус. (внутримозговая опухоль задне-лобной области справа); 5 — частотные спектры ЭЭГ больной В. (арахноидэндотелиома межлолушарной щели в области полюсов лобных долей мозга, больше слева). А — частотные спектры ЭЭГ перед предъявлением индифферентных раздражителей (/), в момент их действия (//) и после их прекращения (///); Б — частотные спектры ЭЭГ перед предъявлением сигнальных раздражителей (/), в момент их действия (счет звуков — //) и после их прекращения (///)
Аналогичные данные, говорящие об участии лобных долей мозга в регуляции состояний активности, были 'получаны в специальном исследовании, проведенном за последнее время Е. Ю. Артемьевой и Е. Д.Хомской.
(Как показали наблюдения А. А. Генкина (1962), электрическая активность мозга характеризуется важным показателем, меняющимся в зависимости от колебаний состояния активности. Этот показатель связан с формой волны альфа-ритма. Тщательные измерения обнаружили, что соотношение длительностей восходящего и нисходящего фронтов альфа-волн за определенный отрезок времени характеризуется определенным индексом асимметрии, который отражает изменение функционального состояния мозга в сторону повышения или понижения уровня бодрствования. При исследовании динамики этого показателя (при t=1 сек) было обнаружено (Е. Ю. Артемьева, Л. Д. Мешалкин и Е. Д. Хомская, 1965), что у здоровых испытуемых наблюдаются ритмические колебания асимметрии альфа-волн со строгим сохранением периодики, величиной в 6—8, чаще 7 сек. Стоит, однако, вызвать у испытуемого состояние напряжения (например, предлагая ему выполнить какие-нибудь задачи, производить в уме разные операции счета и т. п.), чтобы этот равномерный ритм колебаний ломался, отраж ая тем самым состояние активации коры. Прекращение деятельности ведет к восстановлению первоначальной периодичности колебаний асимметрии альфа-волн. Данный параметр подчиняется закономерностям ориентировочных реакций, а именно: ломка периодичности колебаний наблюдается только при первых предъявлениях раздражителей, исчезая по ходу их повторения и возникает вновь после введения сигнального значения раздражителей. Как это было установлено Е. Ю. Артемьевой и Е. Д. Хомской (1966), у больных с поражением лобных долей мозга период колебаний асимметрии альфа-волн уменьшен до 3—5 сек,и при выполнении интеллектуальных заданий, а также при предъявлении несигнальных и сигнальных раздражителей отсутствуют значимые изменения этого показателя. Этот факт служит еще одним доказательством того, что лобные доли мозга принимают существенное участие в 'механизмах регуляции процессов активации, и их поражение ведет к патологии этих процессов.
Рис. 78 (стр. 236) приводит иллюстрации этого положения.

Рис. 78. Изменение асимметрии восходящего и нисходящего фронтов альфа-ритма при различных функциональных состояниях в норме и при поражении лобных долей (по E Ю. Артемьевой и Е. Д. Хомской, 1966) / — Нарушение периодичности колебаний асимметрии альфа-волн во время счета (умножение двухзначных чисел в уме) у здоровых испытуемых. А, Б, В,— различные здоровые испытуемые. Под каждым графиком приведены 3-секундные отрезки ЭЭГ. Вертикальная линия означает начало счета. Записи ЭЭГ до и после начала деятельности визуально неразличимы // — Динамика периодических колебаний асимметрии волн ЭЭГ у больного Ус. (внутри-мозговая опухоль правой задне-лобной области мозга) во время счета в уме (вычитание от 100 по 7); отсутствуют нарушения периодичности колебаний асимметрии альфа-волн (верхний график) или даже появляются относительно упорядоченные колебания асимметрии по сравнению с фоном (нижний график)
Нам осталось упомянуть и последнюю группу фактов, полученных в последнее время Е. Д. Хомской и ее сотрудниками.
Известно, что одним из показателей процесса активации являются вызванные потенциалы, возникающие в соответствующих отделах коры головного мозга. Как показали опыты Э. Г. Симерницкой и Е. Д Хомской (1966), инструкция, направляющая внимание испытуемых на различение вспышек или ожидание укола (например, «следите, какая из вспышек длиннее» или «после десятой вспышки будет укол в руку»), вызывает у нормальных испытуемых изменение временных и амплитудных параметров вызванных потенциалов соответственно в затылочных или сензомоторных отделах мозговой коры. Такой же эффект изменения параметров вызванных ответов под влиянием действия словесной инструкции наблюдается и у большинства больных с поражением задних отделов мозга; однако у больных с поражением лобных отделов мозга закономерные сдвиги параметров вызванных потенциалов под влиянием речевой инструкции отсутствуют или эти изменения носят иной характер, чем в норме. Эти факты, иллюстрация которых приводится на рис. 79, служат еще одним подтверждением высказанного выше положения. Близкие к этому данные были получены и в исследованиях Н. А. Филиппычевой (1966), на которых мы еще остановимся.


Р ис 79 Изменение параметров вызванных ответов (ВО) при введении различных инструкций в норме и при поражении лобных долей мозга (по Э Г Симерницкой иE Д Хомской, 1966) / — Изменения параметров ВО при введении различных инструкций у здоровой испытуемой E Л —введение инструкции «после 8-10-й вспышки будет ток в левую руку», Б— введение инструкции «различайте вспышки по длительности», В — введение инструкции «как только увидите вспышку, нажимайте как можно скорее на кнопку левой рукой» Две верхние кривые — затылочно-теменные отведения, две нижние кривые — центрально-лобные отведения. Сплошная линия — ВО при выполнении задания. Пунктирная и штрих-пунктирная линии — ВО до и после выполнения заданий. При инструкции, адресованной к зрительному анализатору, происходит локальное изменение ВО в теменно-затылочных отведениях (Б). В ответ на другие задания локальные изменения ВО обнаруживаются в центрально-лобных отведениях (А и fi) // — ВО при введении различных инструкций у больного У. (опухоль передних отделов серповидного отростка справа). В отличие от нормального испытуемого специфика речевой инструкции не влияет на характер электрической активности. Обозначения те же.
Изложенные нами факты позволяют прийти к очень существенным выводам.
Лобные доли мозга, интимно связанные двусторонними связями с ретикулярной формацией, играют существенную роль в регуляции состояний активности, осуществляемой при участии речи.
Поражение лобных долей мозга приводит к тому, что эти наиболее высокие формы регуляции состояния активности нарушаются. Эти явления, прослеженные за последние годы на целой группе индикаторов, позволяют сделать существенный шаг к более точному физиологическому анализу той роли, которую играют лобные доли мозга в регуляции высших форм психической деятельности, и делают возможным подойти к анализу нарушений сложных видов движений и действий, которые возникают при поражении лобных долей мозга.
Дата добавления: 2015-10-16; просмотров: 137 | Нарушение авторских прав
| <== предыдущая страница | | | следующая страница ==> |
| Б) Данные о строении и функциях лобных отделов мозга | | | А) О строении произвольных движений и действий |